Fuete Síncrono: Motores Biológicos em Nanotecnologia
 Nas vastas extensões de nossa galáxia, muitos segredos estão ocultos que cientistas de todo o mundo estão tentando encontrar e desvendar. No entanto, não é necessário que algo seja grande para ser misterioso. Uma prova vívida disso é o mundo no nível celular. Muitas das mais diversas formas, estruturas, funções e propósitos das células juntas cumprem uma tarefa comum - manter a vida do corpo. Se você exagera, as células, como as pessoas, têm profissões: carteiros, transmitindo informações entre células e tecidos; guardas de fronteira que identificam e combatem infecções; arquivistas coletando e armazenando informações, etc. Nesta incrível variedade de especialidades, existe uma profissão muito incomum, pelo menos para nós, um motor biológico que gera a força mecânica necessária para o movimento das células.Essas células são particularmente interessantes no contexto da nanotecnologia. Anteriormente, havia um problema na implementação de um nanodispositivo viável baseado em motores biológicos - os motores devem ser integrados em sistemas maiores para que seus movimentos mecânicos possam ser efetivamente conectados a outras unidades moleculares. Cientistas da Universidade de Munique (Alemanha) conseguiram se aproximar da implementação desse conceito. Quais células e unidades moleculares específicas foram usadas na criação do modelo, como o trabalho foi controlado, qual era a tarefa do sistema de trabalho e que resultados ele mostrou? Encontraremos respostas para essas perguntas no relatório do grupo de pesquisa. Vai.
Nas vastas extensões de nossa galáxia, muitos segredos estão ocultos que cientistas de todo o mundo estão tentando encontrar e desvendar. No entanto, não é necessário que algo seja grande para ser misterioso. Uma prova vívida disso é o mundo no nível celular. Muitas das mais diversas formas, estruturas, funções e propósitos das células juntas cumprem uma tarefa comum - manter a vida do corpo. Se você exagera, as células, como as pessoas, têm profissões: carteiros, transmitindo informações entre células e tecidos; guardas de fronteira que identificam e combatem infecções; arquivistas coletando e armazenando informações, etc. Nesta incrível variedade de especialidades, existe uma profissão muito incomum, pelo menos para nós, um motor biológico que gera a força mecânica necessária para o movimento das células.Essas células são particularmente interessantes no contexto da nanotecnologia. Anteriormente, havia um problema na implementação de um nanodispositivo viável baseado em motores biológicos - os motores devem ser integrados em sistemas maiores para que seus movimentos mecânicos possam ser efetivamente conectados a outras unidades moleculares. Cientistas da Universidade de Munique (Alemanha) conseguiram se aproximar da implementação desse conceito. Quais células e unidades moleculares específicas foram usadas na criação do modelo, como o trabalho foi controlado, qual era a tarefa do sistema de trabalho e que resultados ele mostrou? Encontraremos respostas para essas perguntas no relatório do grupo de pesquisa. Vai.Base de estudo
Como mencionado acima, um motor biológico é uma célula que gera a força mecânica necessária para realizar o movimento das células, bem como o transporte intracelular. Esses motores incluem proteínas motoras e complexos de proteínas.Para realizar suas atividades não padronizadas, as proteínas motoras, como qualquer máquina, precisam de combustível. Ácido trifosfórico de adenosina (ATP, C 10 H 16 N 5 O 13 P 3 ) desempenha o seu papel . O ATP é uma fonte universal de energia para todos os processos biológicos nos sistemas vivos.As proteínas motoras funcionam por hidrólise * ATP, que permite às proteínas converter energia química em trabalho mecânico.Hidrólise * é a reação de decomposição de troca entre uma substância dissolvida e um solvente, na qual a decomposição de uma substância (neste caso, ATP) e água ocorre com a formação de novos compostos.
Em particular, as proteínas motoras incluem miosinas, cinesinas e dineínas. As miosinas representam cerca de 40-60% de todas as proteínas musculares, participando do processo de contração muscular.As cinesinas, que se tornaram um meme científico há vários anos, viajam através de microtúbulos (estruturas intracelulares de proteínas), participando dos processos de mitose, meiose e transporte vesicular.Demonstração em vídeo de como a cinesina se move através de um microtúbulo, transformado em um meme. (o original é retirado de The Inner Life of a Cell ; a música sobreposta no vídeo é Stayin 'Alive, Bee Gees, 1977).Os dininos, como as cinesinas, também se movem ao longo dos microtúbulos do citoesqueleto, participando do processo de transferência de carga (vesículas, mitocôndrias, etc.).O interesse em biomotores pelos nanotecnologistas deve-se a vários fatores importantes: nanoescala, biocompatibilidade e a capacidade de usar a engenharia genética para criar biomotores com funções especificadas.Atualmente, existem vários desenvolvimentos baseados nos princípios de ação dos motores biológicos. No entanto, informações sobre a quantidade de trabalho real que um único motor molecular pode fazer ainda não são suficientes para uma implementação completa. Outro quebra-cabeça para os cientistas é a questão de como integrar o bloco de movimento molecular em uma estrutura maior, para que seu movimento direcionado seja efetivamente transformado em um aumento da energia potencial em pontos remotos da estrutura.Obviamente, existem muitos obstáculos, mas isso nunca parou os cientistas. No estudo que estamos examinando hoje, os cientistas descrevem a instalação molecular 1, que permite transferir energia potencial da unidade motora para uma unidade receptora remota, acelerando assim o movimento desta última (imagem nº 1).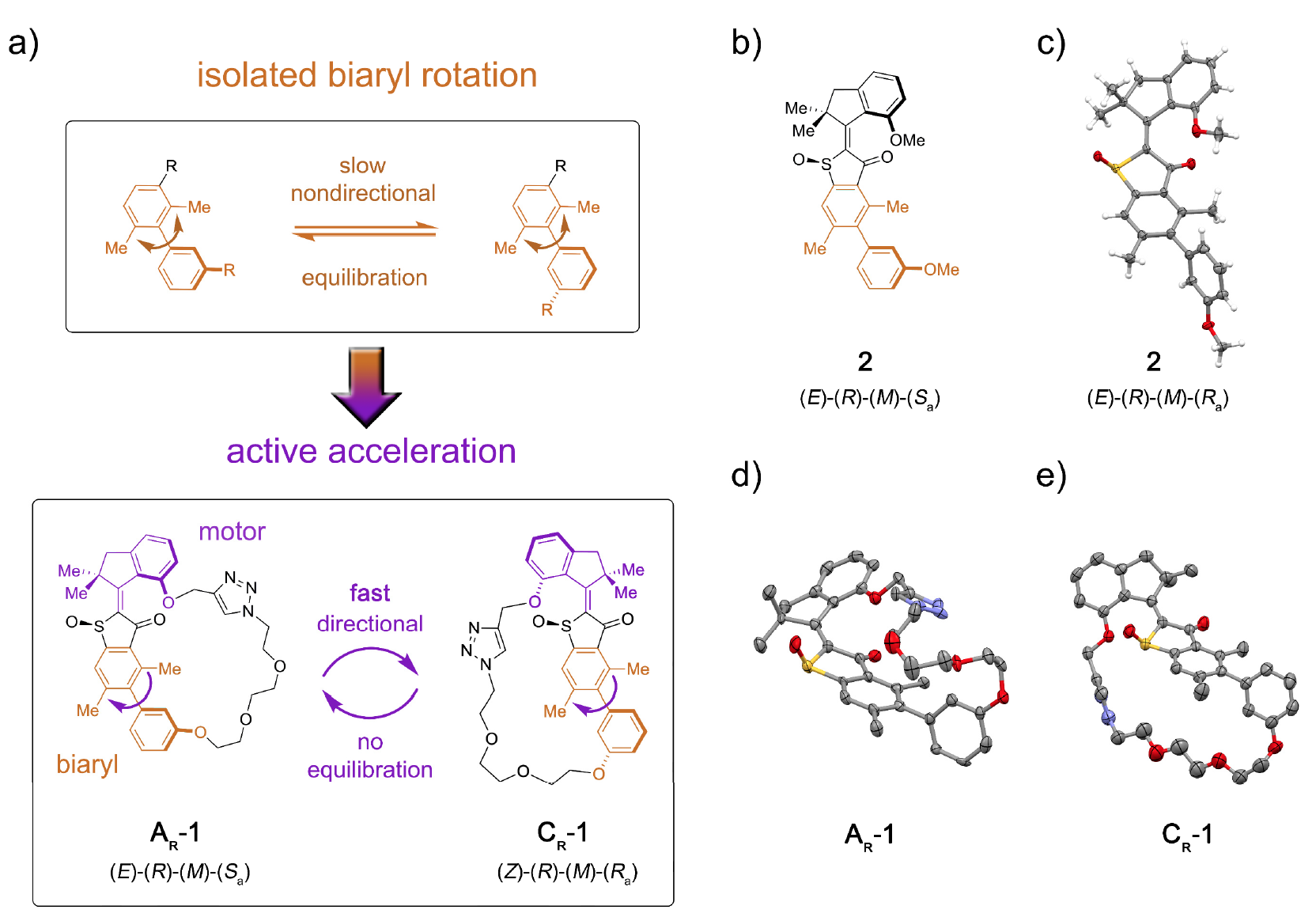 Imagem No. 1O papel da unidade receptora foi desempenhado pelo quiral axial-biaril * , que em sua forma não amarrada (sistema modelo 2) passa por uma rotação lenta e não direcional da atropisomerização na direção do equilíbrio dos enantiômeros 1: 1 ( 1a ).
Imagem No. 1O papel da unidade receptora foi desempenhado pelo quiral axial-biaril * , que em sua forma não amarrada (sistema modelo 2) passa por uma rotação lenta e não direcional da atropisomerização na direção do equilíbrio dos enantiômeros 1: 1 ( 1a ).A quiralidade axial * surge como resultado de um arranjo não-plano de substituintes em relação a algum eixo - o eixo da quiralidade.
Biaril * - Qualquer composto que contenha uma subestrutura, que é uma combinação de dois compostos aromáticos ou grupos aril, se estiverem conectados por uma ligação simples.
A conjugação do bloco motor molecular permite unidirecional desviar essa atropisomerização do equilíbrio. Como resultado, a atropisomerização do biaril não é mais passiva (após a operação do motor), mas representa um obstáculo energético contra o qual o motor deve trabalhar ativamente.Na máquina 1, um aumento na taxa de atropisomerização de biaril em várias ordens de magnitude é alcançado pela ação de um motor que suprime barreiras internas à rotação isolada de biaril (sistema modelo 2 por 1b e 1s ).A Instalação Molecular 1 inclui uma unidade motora molecular baseada no HTI, que pertence à classe dos cromóforos índigo. Este tipo de unidade é extremamente sensível à luz, ou seja, graças à luz, pode ser controlada. O bloco do motor é conectado covalentemente a um eixo biarilo remoto, que não se move com a rotação do motor.Uma das diferenças entre esta instalação e suas versões anteriores é a presença de estados adicionais (etapas): a versão anterior possuía 4 estados e era regulada apenas pelas etapas de rotação do motor; uma nova opção é um sistema de seis velocidades (imagem nº 2).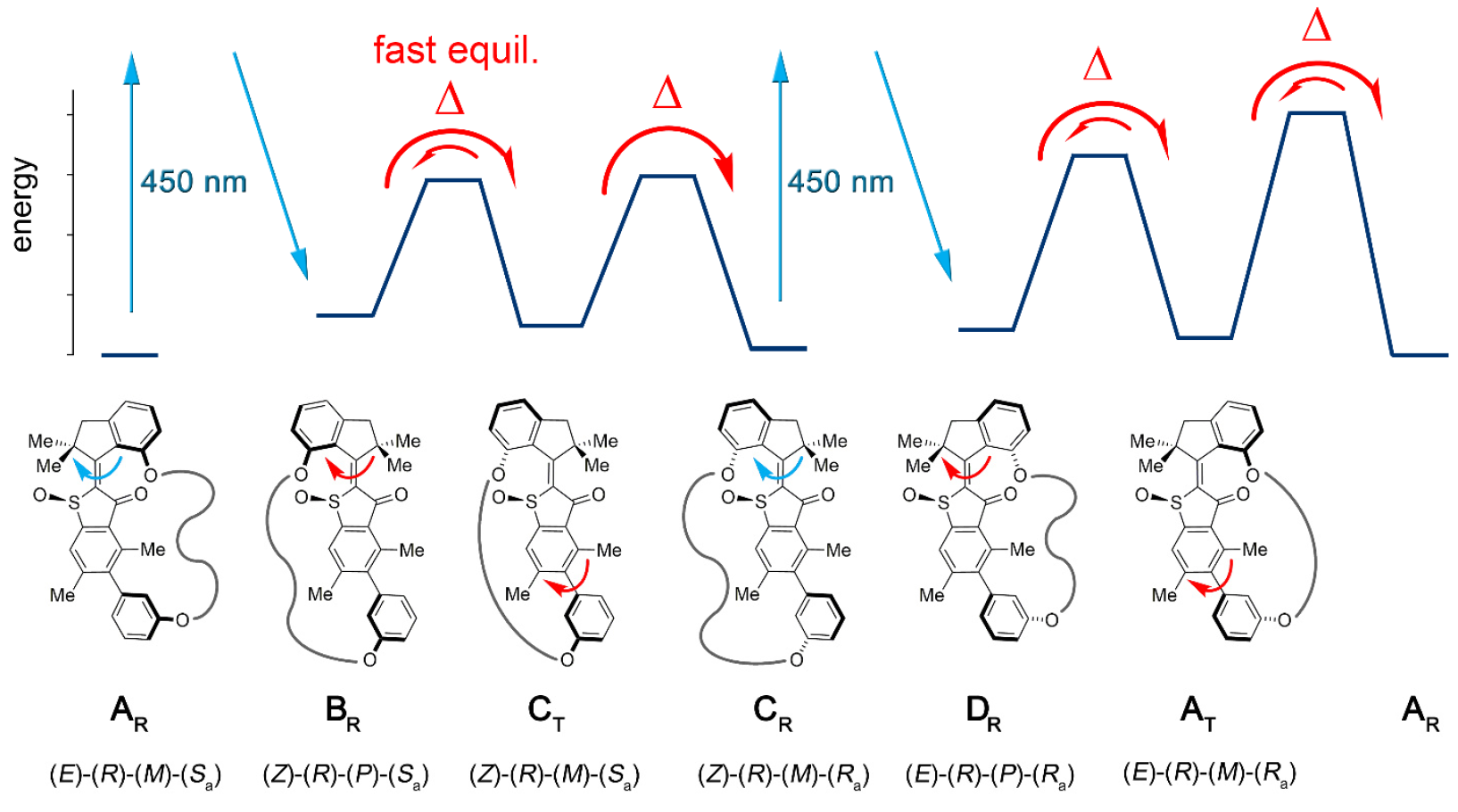 Imagem No. 2Seis estados isoméricos são nomeados da seguinte forma: AT para o estado de estresse A; AR para o estado relaxado A, bem como BR, CT, CR e DR (estados intermediários do sistema).Cinco dessas etapas podem ser observadas experimentalmente, o que confirma a presença da sexta etapa e, portanto, confirma a direcionalidade total para rotação de 360 graus do sistema associado do motor do bloco e do receptor do bloco.O sistema motor 1 foi sintetizado pelo método convergente devido ao precursor de HTI bromado, ao qual uma ligação covalente contendo a função do éster borônico foi acoplada através de uma reação de clique catalisada por cobre.Posteriormente, a reação de Suzuki * fornece macrociclização * , seguida de oxidação para dar a estrutura final 1.
Imagem No. 2Seis estados isoméricos são nomeados da seguinte forma: AT para o estado de estresse A; AR para o estado relaxado A, bem como BR, CT, CR e DR (estados intermediários do sistema).Cinco dessas etapas podem ser observadas experimentalmente, o que confirma a presença da sexta etapa e, portanto, confirma a direcionalidade total para rotação de 360 graus do sistema associado do motor do bloco e do receptor do bloco.O sistema motor 1 foi sintetizado pelo método convergente devido ao precursor de HTI bromado, ao qual uma ligação covalente contendo a função do éster borônico foi acoplada através de uma reação de clique catalisada por cobre.Posteriormente, a reação de Suzuki * fornece macrociclização * , seguida de oxidação para dar a estrutura final 1.A reação de Suzuki * é uma reação orgânica dos ácidos aril e vinilborônico com haletos de aril ou vinila, catalisados por complexos de Pd (0).
Macrociclização * - uma reação de ciclização que leva à formação de um macrociclo, ou seja, um composto químico no qual existem 9 ou mais átomos ligados formando um anel.
Para os dois estados mais estáveis de AR e CR do sistema 1 e o estado mais estável do sistema 2, foram obtidos cristais adequados para análise estrutural (isômeros configurados com ® de racémico * AR em 1d ; CR em 1e ).Racemat * é uma mistura equimolar de dois estereoisômeros, que são imagens espelhadas um do outro. O composto racêmico consiste em cristais, em cada um dos quais existem moléculas de ambos os enantiômeros, e sua proporção é de 1: 1.
Como AR / AT e CR podem ser separados por HPLC (cromatografia líquida de alta eficiência), seu comportamento sob condições de aquecimento e irradiação a diferentes temperaturas pode ser estudado independentemente. Quando a solução de CR racêmica em (CDCl 2 ) 2 foi aquecida a 80 ° C - 140 ° C, AR mais estável foi formado em 93% dos casos. Isso define a diferença de entalpia livre ∆G = 1,8-1,9 kcal / mol nessa faixa de temperatura entre esses dois estados. A análise cinética correspondente revelou uma barreira de alta energia de 28,2 kcal / mol a 80 ° C, acompanhando a isomerização térmica da ligação dupla Z / E.Ao resfriar CR puro a -105 ° C em CD 2 Cl 2 / CS 2(razão 4/1) e irradiando-a com luz de 450 nm, aparece um novo conjunto de sinais, que difere do conjunto conhecido de sinais AR e AT ( 3a ). Imagem No. 3Os sinais do isômero DR decaem para 75% dentro de 28 minutos a uma temperatura de -80 ° C e escuridão completa, mas os sinais do isômero AT, pelo contrário, crescem ( 3 ).O equilíbrio térmico entre DR (os 12% restantes) e AT (88%) é observado a -60 ° C, expresso em expressedG = 0,84 kcal / mol entre os dois estados. A análise cinética da decomposição térmica revelou uma entalpia de ativação livre que acompanha ∆ ‡ G = 13,9 kcal / mol a -80 ° C para este processo. A temperaturas de -40 a 0 ° C, a atenuação dos sinais AT e um aumento concomitante nos sinais RA conhecidos são observados até que o equilíbrio seja alcançado entre eles. Uma análise cinética desse processo determinou equal G igual a 18,4 a 19,3 kcal / mol em temperaturas de –40 ... 0 ° C.Assim, verificou-se que a irradiação do CR leva ao primeiro produto de fotoisomerização DR, que é termicamente convertido no isômero AT, invertendo uma única hélice no bloco motor. Ao resfriar uma solução de equilíbrio de AR / AT no CD 2Cl 2 / CS 2 até -105 ° C e quando foi irradiado com luz de 450 nm, foi observada a fotorreação da AR, devido à sua prevalência. Também aparece um novo conjunto de sinais que difere dos sinais CR ( 3f ) já conhecidos . Esses novos sinais, sendo o produto da fotorreação AR, confirmam a direção esperada da fotoisomerização de AR para BR, que então se estabiliza ainda mais, passando por rápida inversão helicoidal para TC mesmo em baixa temperatura.A uma temperatura de -80 ° C e na escuridão completa, os sinais BR / CT desaparecem quase completamente, e apenas os sinais CR continuam a amplificar ainda mais ( 3e ). Isso fornece um limite mais baixo para a diferença de energia entre CT e CR igual a 0,98 kcal / mol.A análise cinética determinou G = 13,4–13,5 kcal / mol, o que é necessário para a conversão de ST em CR. Pode-se observar que a energia de ativação da atropisomerização da TC na RC diminui em comparação com a da atropisomerização da AT na AR (18,4 kcal / mol). Isso pode ser devido ao maior estresse (tensão) na TC em comparação ao AT, que pode ser devido às grandes distâncias entre os dois átomos de oxigênio fenólico, que servem como pontos de conexão da cadeia ligante nas estruturas C, que não estão nas estruturas A. No estado cristalino, CR entre dois átomos de oxigênio é 10,6, enquanto na RA é 7,6. Isso confere à cadeia de ligação maior liberdade conformacional (posição espacial dos átomos) nas estruturas A do que nas estruturas C. Se considerarmos a entalpia de ativação livre para a transformação térmica de CT em CR,então este processo de conversão pode ser completamente parado a -105 ° C.A uma temperatura de -105 ° C, é possível verificar o grau de acúmulo de TC durante forte exposição à luz RA. Isso é possível se a diferença de energia entre BR e ST exceder 2,0 kcal / mol e se a estrutura ST não estiver envolvida em nenhum processo fotoquímico por si só. No entanto, o acúmulo de TC durante a irradiação com RA a -105 ° C não é possível, pois há um rápido equilíbrio térmico entre CT e BR.
Imagem No. 3Os sinais do isômero DR decaem para 75% dentro de 28 minutos a uma temperatura de -80 ° C e escuridão completa, mas os sinais do isômero AT, pelo contrário, crescem ( 3 ).O equilíbrio térmico entre DR (os 12% restantes) e AT (88%) é observado a -60 ° C, expresso em expressedG = 0,84 kcal / mol entre os dois estados. A análise cinética da decomposição térmica revelou uma entalpia de ativação livre que acompanha ∆ ‡ G = 13,9 kcal / mol a -80 ° C para este processo. A temperaturas de -40 a 0 ° C, a atenuação dos sinais AT e um aumento concomitante nos sinais RA conhecidos são observados até que o equilíbrio seja alcançado entre eles. Uma análise cinética desse processo determinou equal G igual a 18,4 a 19,3 kcal / mol em temperaturas de –40 ... 0 ° C.Assim, verificou-se que a irradiação do CR leva ao primeiro produto de fotoisomerização DR, que é termicamente convertido no isômero AT, invertendo uma única hélice no bloco motor. Ao resfriar uma solução de equilíbrio de AR / AT no CD 2Cl 2 / CS 2 até -105 ° C e quando foi irradiado com luz de 450 nm, foi observada a fotorreação da AR, devido à sua prevalência. Também aparece um novo conjunto de sinais que difere dos sinais CR ( 3f ) já conhecidos . Esses novos sinais, sendo o produto da fotorreação AR, confirmam a direção esperada da fotoisomerização de AR para BR, que então se estabiliza ainda mais, passando por rápida inversão helicoidal para TC mesmo em baixa temperatura.A uma temperatura de -80 ° C e na escuridão completa, os sinais BR / CT desaparecem quase completamente, e apenas os sinais CR continuam a amplificar ainda mais ( 3e ). Isso fornece um limite mais baixo para a diferença de energia entre CT e CR igual a 0,98 kcal / mol.A análise cinética determinou G = 13,4–13,5 kcal / mol, o que é necessário para a conversão de ST em CR. Pode-se observar que a energia de ativação da atropisomerização da TC na RC diminui em comparação com a da atropisomerização da AT na AR (18,4 kcal / mol). Isso pode ser devido ao maior estresse (tensão) na TC em comparação ao AT, que pode ser devido às grandes distâncias entre os dois átomos de oxigênio fenólico, que servem como pontos de conexão da cadeia ligante nas estruturas C, que não estão nas estruturas A. No estado cristalino, CR entre dois átomos de oxigênio é 10,6, enquanto na RA é 7,6. Isso confere à cadeia de ligação maior liberdade conformacional (posição espacial dos átomos) nas estruturas A do que nas estruturas C. Se considerarmos a entalpia de ativação livre para a transformação térmica de CT em CR,então este processo de conversão pode ser completamente parado a -105 ° C.A uma temperatura de -105 ° C, é possível verificar o grau de acúmulo de TC durante forte exposição à luz RA. Isso é possível se a diferença de energia entre BR e ST exceder 2,0 kcal / mol e se a estrutura ST não estiver envolvida em nenhum processo fotoquímico por si só. No entanto, o acúmulo de TC durante a irradiação com RA a -105 ° C não é possível, pois há um rápido equilíbrio térmico entre CT e BR. Tabela Nº 1A combinação das observações e dados acima permite criar uma imagem mecânica de uma sequência de seis etapas de rotação unidirecional no modelo 1 (imagens nº 2 e nº 4).
Tabela Nº 1A combinação das observações e dados acima permite criar uma imagem mecânica de uma sequência de seis etapas de rotação unidirecional no modelo 1 (imagens nº 2 e nº 4). Imagem No. 4Experimentos com diferentes condições de temperatura nos permitiram determinar a dependência de temperatura de ∆G entre CR e AR / AT, DR e AT, AT e AR, bem como a dependência de temperatura de ∆ G para BR / CT e CR, DR e AT, AT e AR (tabela No. 1)Diferentemente do mecanismo usual de 4 velocidades, baseado no motor HTI, a rotação completa de 360 graus inclui duas etapas adicionais, que são o aumento da energia potencial transmitida do motor ao bloco biarilo. Durante estas duas etapas, uma ligação de energia é formada entre o motor e o bloco biaril ( 4b ), isto é, o último não é mais degenerado energeticamente.
Imagem No. 4Experimentos com diferentes condições de temperatura nos permitiram determinar a dependência de temperatura de ∆G entre CR e AR / AT, DR e AT, AT e AR, bem como a dependência de temperatura de ∆ G para BR / CT e CR, DR e AT, AT e AR (tabela No. 1)Diferentemente do mecanismo usual de 4 velocidades, baseado no motor HTI, a rotação completa de 360 graus inclui duas etapas adicionais, que são o aumento da energia potencial transmitida do motor ao bloco biarilo. Durante estas duas etapas, uma ligação de energia é formada entre o motor e o bloco biaril ( 4b ), isto é, o último não é mais degenerado energeticamente. Imagem nº 5Em vez disso, a energia de qualquer um dos atropisômeros aumenta devido a etapas “catraca” do motor durante uma rotação completa ( 5se 5d ). Essa mudança de energia fornece até 90% da interconversão do atropisômero nos seguintes estágios termicamente ativados e, portanto, rotação quase completa do bloco biaril durante um ciclo de trabalho do mecanismo.Esta observação pode indicar que o bloco biaril sofreu um aumento na energia potencial, que é o resultado da operação do bloco motor. Vale ressaltar que o aumento inicial de energia é muito maior do que o que atinge o bloco biaril. Assim, apenas 72% é transmitido do motor para o bloco biaril.As observações acima sugerem que o comprimento da conexão entre o motor e o bloco biaril é um aspecto importante no processo de alteração do grau de transferência de energia. Portanto, uma conexão mais curta pode permitir que mais energia potencial seja transferida de BR e DR para AT e CT.Para um conhecimento mais detalhado das nuances do estudo, recomendo que você analise o relatório dos cientistas e materiais adicionais .
Imagem nº 5Em vez disso, a energia de qualquer um dos atropisômeros aumenta devido a etapas “catraca” do motor durante uma rotação completa ( 5se 5d ). Essa mudança de energia fornece até 90% da interconversão do atropisômero nos seguintes estágios termicamente ativados e, portanto, rotação quase completa do bloco biaril durante um ciclo de trabalho do mecanismo.Esta observação pode indicar que o bloco biaril sofreu um aumento na energia potencial, que é o resultado da operação do bloco motor. Vale ressaltar que o aumento inicial de energia é muito maior do que o que atinge o bloco biaril. Assim, apenas 72% é transmitido do motor para o bloco biaril.As observações acima sugerem que o comprimento da conexão entre o motor e o bloco biaril é um aspecto importante no processo de alteração do grau de transferência de energia. Portanto, uma conexão mais curta pode permitir que mais energia potencial seja transferida de BR e DR para AT e CT.Para um conhecimento mais detalhado das nuances do estudo, recomendo que você analise o relatório dos cientistas e materiais adicionais .Epílogo
Neste trabalho, os cientistas descreveram o princípio de operação de sua invenção, com base na interação de motores biológicos e elementos moleculares. A instalação desenvolvida permite transferir energia potencial do motor para um elemento específico (neste caso, é um bloco biarilo). Como resultado da atividade motora, a atropisomerização térmica do biaril é forçada a prosseguir unidirecionalmente e a uma taxa mais rápida. A transferência de energia ocorre durante catracas termicamente ativadas (etapas) no mecanismo de rotação. Verificou-se também que cerca de 72% da energia inicial do motor é transmitida para uma determinada unidade.Esta invenção é um passo importante para a criação de dispositivos nanométricos biológicos completos. O uso de biomotores no projeto de tais mecanismos deve-se à sua biocompatibilidade, facilidade de alteração de funcionalidade devido à engenharia genética e à nanoescala natural. No momento, os motores artificiais não podem se orgulhar da combinação de vantagens acima.Os pesquisadores dizem que o próximo passo em seu trabalho será demonstrar que sua instalação é capaz de executar não apenas ações abstratas, mas também ser útil. Em outras palavras, sua criação futura será mais refinada para desempenhar funções úteis no nível molecular.Obrigado pela atenção, continuem curiosos e tenham uma boa semana de trabalho, pessoal. :)Um pouco de publicidade :)
Obrigado por ficar com a gente. Você gosta dos nossos artigos? Deseja ver materiais mais interessantes? Ajude-nos fazendo um pedido ou recomendando aos seus amigos o VPS na nuvem para desenvolvedores a partir de US $ 4,99 , um analógico exclusivo de servidores de nível básico que foi inventado por nós para você: Toda a verdade sobre o VPS (KVM) E5-2697 v3 (6 núcleos) 10 GB DDR4 480 GB SSD 1 Gbps de US $ 19 ou como dividir o servidor? (as opções estão disponíveis com RAID1 e RAID10, até 24 núcleos e até 40GB DDR4).Dell R730xd 2 vezes mais barato no data center Equinix Tier IV em Amsterdã? Somente nós temos 2 TVs Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV a partir de US $ 199 na Holanda!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - a partir de US $ 99! Leia sobre Como criar um prédio de infraestrutura. classe c usando servidores Dell R730xd E5-2650 v4 que custam 9.000 euros por um centavo?Source: https://habr.com/ru/post/undefined/
All Articles