Fuete Sinkron: Motor Biologis dalam Nanoteknologi
 Dalam bentangan luas galaksi kita, banyak rahasia tersembunyi yang para ilmuwan dari seluruh dunia berusaha keras untuk menemukan dan mengungkapnya. Namun, tidak perlu sesuatu menjadi besar untuk menjadi misterius. Bukti nyata dari hal ini adalah dunia berada pada level seluler. Banyak dari bentuk, struktur, fungsi, dan tujuan sel yang paling beragam secara bersamaan memenuhi tugas bersama - menjaga kehidupan tubuh. Jika Anda melebih-lebihkan, maka sel, seperti orang, memiliki profesi: tukang pos, mentransmisikan informasi antara sel dan jaringan; penjaga perbatasan yang mengidentifikasi dan melawan infeksi; arsiparis mengumpulkan dan menyimpan informasi, dll. Dalam rentang spesialisasi yang luar biasa ini, ada profesi yang sangat luar biasa, setidaknya bagi kita, sebuah motor biologis, yang menghasilkan kekuatan mekanis yang diperlukan untuk pergerakan sel.Sel-sel ini sangat menarik dalam konteks nanoteknologi. Sebelumnya, ada masalah dalam implementasi perangkat nano yang bisa dikerjakan berdasarkan pada motor biologis - motor harus diintegrasikan ke dalam sistem yang lebih besar sehingga gerakan mekanis mereka dapat dihubungkan secara efektif dengan unit molekul lainnya. Para ilmuwan dari University of Munich (Jerman) berhasil mendekati implementasi konsep ini. Apa sel spesifik dan unit molekuler yang digunakan dalam menciptakan model, bagaimana pekerjaan mereka dikontrol, apa tugas sistem kerja dan apa hasil yang ditunjukkannya? Kami akan menemukan jawaban untuk pertanyaan-pertanyaan ini dalam laporan kelompok penelitian. Pergilah.
Dalam bentangan luas galaksi kita, banyak rahasia tersembunyi yang para ilmuwan dari seluruh dunia berusaha keras untuk menemukan dan mengungkapnya. Namun, tidak perlu sesuatu menjadi besar untuk menjadi misterius. Bukti nyata dari hal ini adalah dunia berada pada level seluler. Banyak dari bentuk, struktur, fungsi, dan tujuan sel yang paling beragam secara bersamaan memenuhi tugas bersama - menjaga kehidupan tubuh. Jika Anda melebih-lebihkan, maka sel, seperti orang, memiliki profesi: tukang pos, mentransmisikan informasi antara sel dan jaringan; penjaga perbatasan yang mengidentifikasi dan melawan infeksi; arsiparis mengumpulkan dan menyimpan informasi, dll. Dalam rentang spesialisasi yang luar biasa ini, ada profesi yang sangat luar biasa, setidaknya bagi kita, sebuah motor biologis, yang menghasilkan kekuatan mekanis yang diperlukan untuk pergerakan sel.Sel-sel ini sangat menarik dalam konteks nanoteknologi. Sebelumnya, ada masalah dalam implementasi perangkat nano yang bisa dikerjakan berdasarkan pada motor biologis - motor harus diintegrasikan ke dalam sistem yang lebih besar sehingga gerakan mekanis mereka dapat dihubungkan secara efektif dengan unit molekul lainnya. Para ilmuwan dari University of Munich (Jerman) berhasil mendekati implementasi konsep ini. Apa sel spesifik dan unit molekuler yang digunakan dalam menciptakan model, bagaimana pekerjaan mereka dikontrol, apa tugas sistem kerja dan apa hasil yang ditunjukkannya? Kami akan menemukan jawaban untuk pertanyaan-pertanyaan ini dalam laporan kelompok penelitian. Pergilah.Dasar studi
Seperti disebutkan di atas, motor biologis adalah sel yang menghasilkan kekuatan mekanis yang diperlukan untuk mewujudkan pergerakan sel, serta transportasi intraseluler. Motor ini termasuk protein motor dan kompleks protein.Untuk melakukan aktivitasnya yang tidak standar, protein motor, seperti mesin apa pun, membutuhkan bahan bakar. Asam adenosin trifosfat (ATP, C 10 H 16 N 5 O 13 P 3 ) memainkan perannya . ATP adalah sumber energi universal untuk semua proses biologis dalam sistem kehidupan.Protein motor berfungsi dengan hidrolisis * ATP, yang memungkinkan protein mengubah energi kimia menjadi kerja mekanis.Hidrolisis * adalah reaksi pertukaran dekomposisi antara zat terlarut dan pelarut, di mana dekomposisi suatu zat (dalam hal ini ATP) dan air terjadi dengan pembentukan senyawa baru.
Secara khusus, protein motorik termasuk miosin, kinesin dan dynein. Myosins membentuk sekitar 40-60% dari semua protein otot, berpartisipasi dalam proses kontraksi otot.Kinesin, yang menjadi meme ilmiah beberapa tahun yang lalu, melakukan perjalanan melalui mikrotubulus (struktur intraseluler protein), berpartisipasi dalam proses mitosis, meiosis dan transportasi vesikuler.Video demonstrasi tentang bagaimana kinesin bergerak melalui mikrotubulus, diubah menjadi meme. (aslinya diambil dari The Inner Life of a Cell ; musik yang ditayangkan di video adalah Stayin 'Alive, Bee Gees, 1977).Dynein, seperti kinesin, juga bergerak sepanjang mikrotubulus sitoskeleton, berpartisipasi dalam proses pemindahan kargo (vesikel, mitokondria, dll.).Ketertarikan pada biomotor oleh para nanoteknologi disebabkan oleh beberapa faktor penting: skala nano, biokompatibilitas dan kemampuan untuk menggunakan rekayasa genetika untuk membuat biomotor dengan fungsi-fungsi tertentu.Saat ini, ada sejumlah perkembangan berdasarkan prinsip aksi motor biologis. Namun, informasi mengenai seberapa banyak kerja nyata yang dapat dilakukan oleh motor molekuler tunggal belum cukup untuk implementasi penuh. Teka-teki lain bagi para ilmuwan adalah pertanyaan tentang bagaimana mengintegrasikan blok gerak molekuler ke dalam struktur yang lebih besar sehingga gerakan terarahnya diubah secara efektif menjadi peningkatan energi potensial pada titik-titik terpencil dalam struktur.Jelas, ada banyak kendala, tetapi ini tidak pernah menghentikan para ilmuwan. Dalam studi yang kami pertimbangkan saat ini, para ilmuwan menggambarkan instalasi molekul 1, yang memungkinkan transfer energi potensial dari unit motor ke unit penerima jarak jauh, sehingga mempercepat pergerakan yang terakhir (gambar No. 1).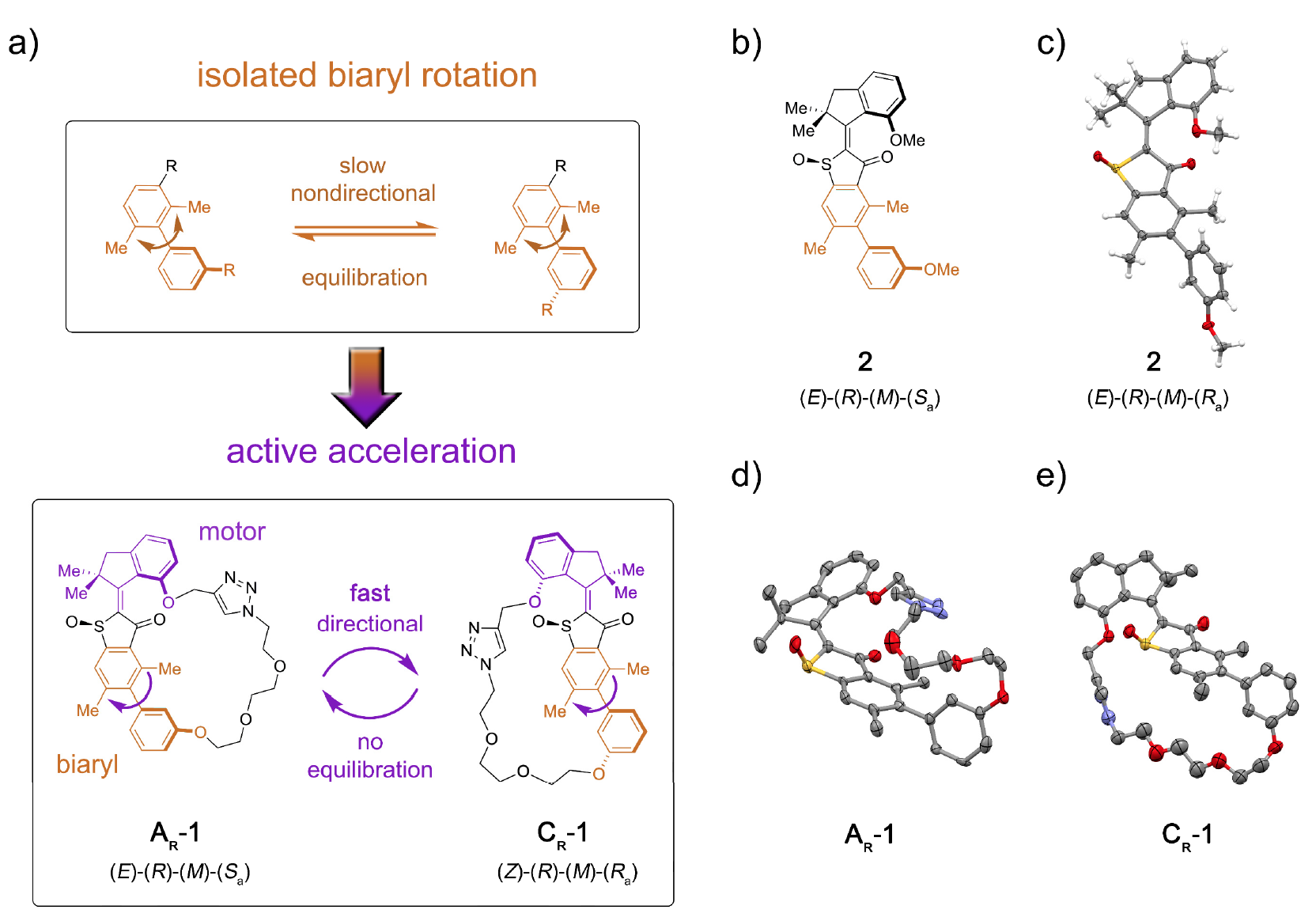 Gambar No. 1Peran unit penerima dimainkan oleh axial-chiral * biaryl * , yang dalam bentuk non-tethered (sistem model 2) mengalami rotasi atropisomerisasi yang lambat dan tidak terarah ke arah keseimbangan enansiomer 1: 1 ( 1a ).
Gambar No. 1Peran unit penerima dimainkan oleh axial-chiral * biaryl * , yang dalam bentuk non-tethered (sistem model 2) mengalami rotasi atropisomerisasi yang lambat dan tidak terarah ke arah keseimbangan enansiomer 1: 1 ( 1a ).Chirality aksial * muncul sebagai akibat dari pengaturan substitusi nonplanar relatif terhadap beberapa sumbu - sumbu chirality.
Biaryl * - Setiap senyawa yang mengandung substruktur, yang merupakan kombinasi dari dua senyawa aromatik atau kelompok aril, jika mereka dihubungkan oleh ikatan tunggal.
Konjugasi dari blok motor molekuler memungkinkan searah untuk mengalihkan atropisomerisasi ini dari kesetimbangan. Akibatnya, atropisomerisasi biaryl tidak lagi pasif (mengikuti operasi motor), tetapi merupakan hambatan energi terhadap mana motor harus aktif bekerja.Dalam mesin 1, peningkatan laju atropisomerisasi biaryl oleh beberapa urutan besarnya dicapai dengan aksi motor yang menekan hambatan internal ke rotasi biaryl yang terisolasi (sistem model 2 oleh 1b dan 1s ).Instalasi Molekuler 1 termasuk unit motor molekuler berdasarkan HTI, yang termasuk dalam kelas kromofor indigo. Jenis unit ini sangat sensitif terhadap cahaya, mis. berkat cahayanya, bisa dikendalikan. Blok motor secara kovalen terhubung ke sumbu biaryl jarak jauh, yang tidak bergerak dengan putaran motor.Salah satu perbedaan antara instalasi ini dan versi sebelumnya adalah adanya status tambahan (langkah-langkah): versi sebelumnya memiliki 4 status dan hanya diatur oleh langkah-langkah rotasi motor; opsi baru adalah sistem enam kecepatan (gambar No. 2).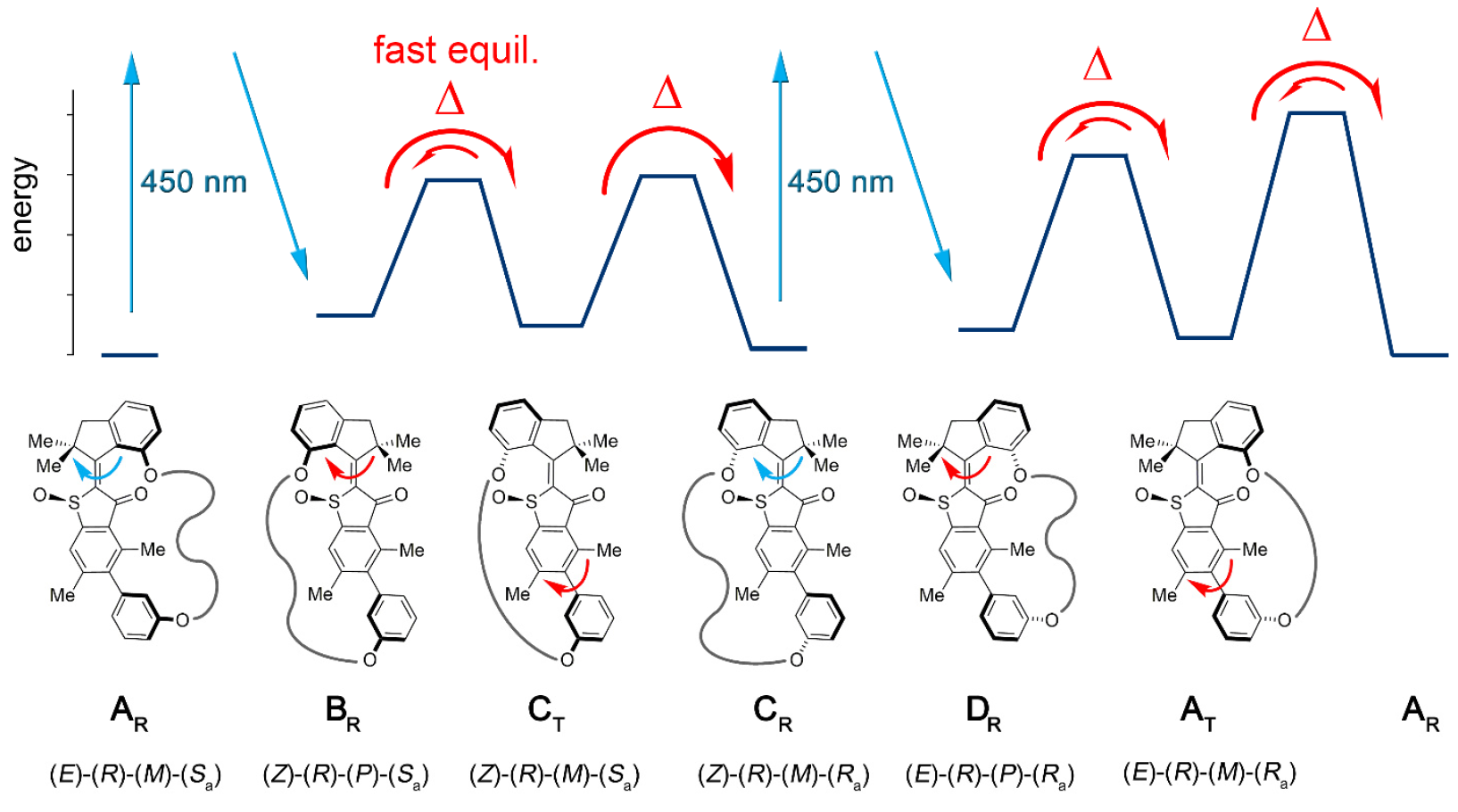 Gambar No. 2Enam keadaan isomer dinamai sebagai berikut: AT untuk keadaan tegangan A; AR untuk keadaan santai A, serta BR, CT, CR, dan DR (kondisi tengah sistem).Lima dari langkah-langkah ini dapat diamati secara eksperimental, yang mengkonfirmasi keberadaan langkah keenam dan, oleh karena itu, mengkonfirmasi arah penuh untuk rotasi 360 derajat dari sistem yang terkait dari motor blok dan penerima blok.Sistem motor 1 disintesis oleh metode konvergen karena prekursor HTI brominasi, dimana ikatan kovalen yang mengandung fungsi ester boronat dilekatkan melalui reaksi klik yang dikatalisis tembaga.Selanjutnya, reaksi Suzuki * memberikan macrocyclization * , diikuti oleh oksidasi untuk memberikan struktur akhir 1.
Gambar No. 2Enam keadaan isomer dinamai sebagai berikut: AT untuk keadaan tegangan A; AR untuk keadaan santai A, serta BR, CT, CR, dan DR (kondisi tengah sistem).Lima dari langkah-langkah ini dapat diamati secara eksperimental, yang mengkonfirmasi keberadaan langkah keenam dan, oleh karena itu, mengkonfirmasi arah penuh untuk rotasi 360 derajat dari sistem yang terkait dari motor blok dan penerima blok.Sistem motor 1 disintesis oleh metode konvergen karena prekursor HTI brominasi, dimana ikatan kovalen yang mengandung fungsi ester boronat dilekatkan melalui reaksi klik yang dikatalisis tembaga.Selanjutnya, reaksi Suzuki * memberikan macrocyclization * , diikuti oleh oksidasi untuk memberikan struktur akhir 1.Reaksi Suzuki * adalah reaksi organik asam aril dan vinilboronat dengan aril atau vinil halida, dikatalisis oleh kompleks Pd (0).
Macrocyclization * - reaksi siklisasi yang mengarah pada pembentukan macrocycle, mis. suatu senyawa kimia yang mengandung 9 atom atau lebih yang membentuk cincin.
Untuk dua keadaan paling stabil dari AR dan CR dari sistem 1 dan keadaan paling stabil dari sistem 2, kristal yang cocok untuk analisis struktural diperoleh (isomer-isomer rasemik * AR yang terkonfigurasi pada 1d ; CR pada 1e ).Racemat * adalah campuran equimolar dari dua stereoisomer, yang merupakan gambar cermin satu sama lain. Senyawa rasemik terdiri dari kristal, di mana masing-masing terdapat molekul kedua enansiomer, dan perbandingannya adalah 1: 1.
Karena AR / AT dan CR dapat dipisahkan menggunakan HPLC (kromatografi cair kinerja tinggi), perilaku mereka dalam kondisi pemanasan dan iradiasi pada suhu yang berbeda dapat dipelajari secara independen. Ketika solusi CR rasematik dalam (CDCl 2 ) 2 dipanaskan hingga 80 ° C - 140 ° C, AR yang lebih stabil terbentuk pada 93% kasus. Ini menetapkan perbedaan dalam entalpi bebas ∆G = 1,8–1,9 kkal / mol dalam kisaran suhu ini di antara kedua kondisi ini. Analisis kinetik yang sesuai mengungkapkan penghalang energi tinggi 28,2 kkal / mol pada 80 ° C, yang menyertai isomerisasi termal ikatan rangkap Z / E.Saat mendinginkan CR murni hingga -105 ° C dalam CD 2 Cl 2 / CS 2(rasio 4/1) dan irradiasinya dengan cahaya 450 nm, satu set sinyal baru muncul, yang berbeda dari set sinyal AR dan AT ( 3a ) yang diketahui . Gambar No. 3Sinyal-sinyal dari DR isomer membusuk menjadi 75% dalam waktu 28 menit pada suhu -80 ° C dan gelap gulita, tetapi sinyal-sinyal dari isomer AT, sebaliknya, tumbuh ( 3 ).Kesetimbangan termal antara DR (12% sisanya) dan AT (88%) diamati pada -60 ° C, yang dinyatakan dalam ∆G = 0,84 kkal / mol antara kedua kondisi. Analisis kinetik dari dekomposisi termal mengungkapkan entalpi aktivasi bebas yang menyertai ∆ ‡ G = 13,9 kkal / mol pada -80 ° C untuk proses ini. Pada suhu dari -40 hingga 0 ° C, pelemahan sinyal AT dan peningkatan secara bersamaan sinyal AR yang diketahui diamati sampai tercapai keseimbangan di antara keduanya. Analisis kinetik dari proses ini menentukan ∆ ‡ G sama dengan dari 18,4 hingga 19,3 kkal / mol pada suhu dari -40 ... 0 ° CDengan demikian, ditemukan bahwa iradiasi CR mengarah ke produk photoisomerisasi DR pertama, yang secara termal dikonversi ke isomer AT dengan membalikkan heliks tunggal pada blok motor. Saat mendinginkan solusi kesetimbangan AR / AT dalam CD 2Cl 2 / CS 2 hingga -105 ° C dan ketika diiradiasi dengan cahaya 450 nm, fotoreaksi AR diamati, karena prevalensinya. Seperangkat sinyal baru juga muncul yang berbeda dari sinyal CR ( 3f ) yang sudah diketahui . Sinyal-sinyal baru ini, sebagai produk fotoreaksi AR, mengkonfirmasi arah photoisomerisasi yang diharapkan dari AR ke BR, yang kemudian semakin stabil, menjalani inversi heliks cepat ke CT bahkan pada suhu rendah.Pada suhu -80 ° C dan dalam kegelapan total, sinyal BR / CT hampir sepenuhnya menghilang, dan hanya sinyal CR yang terus menguat lebih lanjut ( 3e ). Ini memberikan batas yang lebih rendah untuk perbedaan energi antara CT dan CR sebesar 0,98 kkal / mol.Analisis kinetik ditentukan ∆ ‡ G = 13,4-13,5 kkal / mol, yang diperlukan untuk konversi ST menjadi CR. Dapat dilihat bahwa energi aktivasi atropisomerisasi CT dalam CR menurun dibandingkan dengan yang untuk atropisomerisasi AT dalam AR (18,4 kkal / mol). Hal ini mungkin disebabkan oleh tekanan (regangan) yang lebih tinggi pada CT dibandingkan dengan AT, yang mungkin disebabkan oleh jarak yang besar antara dua atom oksigen fenolik, yang berfungsi sebagai titik koneksi rantai penghubung pada struktur C, yang tidak pada struktur A. Pada kondisi kristal, CR antara dua atom oksigen adalah 10.6, sedangkan di AR adalah 7.6. Ini memberi rantai penghubung kebebasan konformasi (posisi spasial atom) yang lebih besar dalam struktur-A daripada struktur-C. Jika kita memperhitungkan entalpi aktivasi gratis untuk transformasi termal CT menjadi CR,maka proses konversi ini dapat sepenuhnya dihentikan pada -105 ° C.Pada suhu -105 ° C, dimungkinkan untuk memeriksa tingkat akumulasi CT selama paparan cahaya AR yang kuat. Ini dimungkinkan jika perbedaan energi antara BR dan ST melebihi 2.0 kkal / mol dan jika struktur ST tidak terlibat dalam proses fotokimia dengan sendirinya. Namun, akumulasi CT selama iradiasi AR pada -105 ° C tidak dimungkinkan, karena ada keseimbangan termal yang cepat antara CT dan BR.
Gambar No. 3Sinyal-sinyal dari DR isomer membusuk menjadi 75% dalam waktu 28 menit pada suhu -80 ° C dan gelap gulita, tetapi sinyal-sinyal dari isomer AT, sebaliknya, tumbuh ( 3 ).Kesetimbangan termal antara DR (12% sisanya) dan AT (88%) diamati pada -60 ° C, yang dinyatakan dalam ∆G = 0,84 kkal / mol antara kedua kondisi. Analisis kinetik dari dekomposisi termal mengungkapkan entalpi aktivasi bebas yang menyertai ∆ ‡ G = 13,9 kkal / mol pada -80 ° C untuk proses ini. Pada suhu dari -40 hingga 0 ° C, pelemahan sinyal AT dan peningkatan secara bersamaan sinyal AR yang diketahui diamati sampai tercapai keseimbangan di antara keduanya. Analisis kinetik dari proses ini menentukan ∆ ‡ G sama dengan dari 18,4 hingga 19,3 kkal / mol pada suhu dari -40 ... 0 ° CDengan demikian, ditemukan bahwa iradiasi CR mengarah ke produk photoisomerisasi DR pertama, yang secara termal dikonversi ke isomer AT dengan membalikkan heliks tunggal pada blok motor. Saat mendinginkan solusi kesetimbangan AR / AT dalam CD 2Cl 2 / CS 2 hingga -105 ° C dan ketika diiradiasi dengan cahaya 450 nm, fotoreaksi AR diamati, karena prevalensinya. Seperangkat sinyal baru juga muncul yang berbeda dari sinyal CR ( 3f ) yang sudah diketahui . Sinyal-sinyal baru ini, sebagai produk fotoreaksi AR, mengkonfirmasi arah photoisomerisasi yang diharapkan dari AR ke BR, yang kemudian semakin stabil, menjalani inversi heliks cepat ke CT bahkan pada suhu rendah.Pada suhu -80 ° C dan dalam kegelapan total, sinyal BR / CT hampir sepenuhnya menghilang, dan hanya sinyal CR yang terus menguat lebih lanjut ( 3e ). Ini memberikan batas yang lebih rendah untuk perbedaan energi antara CT dan CR sebesar 0,98 kkal / mol.Analisis kinetik ditentukan ∆ ‡ G = 13,4-13,5 kkal / mol, yang diperlukan untuk konversi ST menjadi CR. Dapat dilihat bahwa energi aktivasi atropisomerisasi CT dalam CR menurun dibandingkan dengan yang untuk atropisomerisasi AT dalam AR (18,4 kkal / mol). Hal ini mungkin disebabkan oleh tekanan (regangan) yang lebih tinggi pada CT dibandingkan dengan AT, yang mungkin disebabkan oleh jarak yang besar antara dua atom oksigen fenolik, yang berfungsi sebagai titik koneksi rantai penghubung pada struktur C, yang tidak pada struktur A. Pada kondisi kristal, CR antara dua atom oksigen adalah 10.6, sedangkan di AR adalah 7.6. Ini memberi rantai penghubung kebebasan konformasi (posisi spasial atom) yang lebih besar dalam struktur-A daripada struktur-C. Jika kita memperhitungkan entalpi aktivasi gratis untuk transformasi termal CT menjadi CR,maka proses konversi ini dapat sepenuhnya dihentikan pada -105 ° C.Pada suhu -105 ° C, dimungkinkan untuk memeriksa tingkat akumulasi CT selama paparan cahaya AR yang kuat. Ini dimungkinkan jika perbedaan energi antara BR dan ST melebihi 2.0 kkal / mol dan jika struktur ST tidak terlibat dalam proses fotokimia dengan sendirinya. Namun, akumulasi CT selama iradiasi AR pada -105 ° C tidak dimungkinkan, karena ada keseimbangan termal yang cepat antara CT dan BR. Tabel No. 1Kombinasi dari pengamatan dan data di atas memungkinkan Anda untuk membuat gambar mekanis dari urutan 6 langkah rotasi searah dalam model 1 (gambar No. 2 dan No. 4).
Tabel No. 1Kombinasi dari pengamatan dan data di atas memungkinkan Anda untuk membuat gambar mekanis dari urutan 6 langkah rotasi searah dalam model 1 (gambar No. 2 dan No. 4). Gambar No. 4Eksperimen dengan kondisi suhu yang berbeda memungkinkan kita untuk menentukan ketergantungan suhu ∆G antara CR dan AR / AT, DR dan AT, AT dan AR, serta ketergantungan suhu ∆ G untuk BR / CT dan CR, DR dan AT, AT dan AR (tabel no. 1).Tidak seperti mekanisme 4 langkah biasa yang didasarkan pada motor HTI, rotasi 360 derajat penuh mencakup dua langkah tambahan, yang merupakan peningkatan energi potensial yang ditransmisikan dari motor ke blok biaryl. Selama dua langkah ini, ikatan energi terbentuk antara motor dan blok biaryl ( 4b ), yaitu yang terakhir tidak lagi mengalami degenerasi energi.
Gambar No. 4Eksperimen dengan kondisi suhu yang berbeda memungkinkan kita untuk menentukan ketergantungan suhu ∆G antara CR dan AR / AT, DR dan AT, AT dan AR, serta ketergantungan suhu ∆ G untuk BR / CT dan CR, DR dan AT, AT dan AR (tabel no. 1).Tidak seperti mekanisme 4 langkah biasa yang didasarkan pada motor HTI, rotasi 360 derajat penuh mencakup dua langkah tambahan, yang merupakan peningkatan energi potensial yang ditransmisikan dari motor ke blok biaryl. Selama dua langkah ini, ikatan energi terbentuk antara motor dan blok biaryl ( 4b ), yaitu yang terakhir tidak lagi mengalami degenerasi energi. Image No. 5Sebaliknya, energi dari salah satu atropisomer meningkat karena langkah "ratchet" motor selama satu putaran penuh ( 5sdan 5d ). Perubahan energi ini menyediakan hingga 90% dari interkonversi atropisomer pada tahap-tahap yang diaktifkan secara termal berikut dan, oleh karena itu, rotasi hampir sempurna dari blok biaryl selama satu siklus kerja mekanisme.Pengamatan ini dapat menunjukkan bahwa blok biaryl telah mengalami peningkatan energi potensial, yang merupakan hasil dari operasi blok motorik. Perlu dicatat bahwa peningkatan energi awal jauh lebih besar daripada apa yang mencapai blok biaryl. Jadi, hanya 72% darinya yang ditransmisikan dari motor ke blok biaryl.Pengamatan di atas menunjukkan bahwa panjang hubungan antara motor dan blok biaryl merupakan aspek penting dalam proses mengubah tingkat transfer energi. Oleh karena itu, koneksi yang lebih pendek memungkinkan lebih banyak energi potensial ditransfer dari BR dan DR ke AT dan CT.Untuk seorang kenalan yang lebih mendetail dengan nuansa penelitian ini, saya sarankan Anda melihat laporan para ilmuwan dan bahan tambahan untuk itu.
Image No. 5Sebaliknya, energi dari salah satu atropisomer meningkat karena langkah "ratchet" motor selama satu putaran penuh ( 5sdan 5d ). Perubahan energi ini menyediakan hingga 90% dari interkonversi atropisomer pada tahap-tahap yang diaktifkan secara termal berikut dan, oleh karena itu, rotasi hampir sempurna dari blok biaryl selama satu siklus kerja mekanisme.Pengamatan ini dapat menunjukkan bahwa blok biaryl telah mengalami peningkatan energi potensial, yang merupakan hasil dari operasi blok motorik. Perlu dicatat bahwa peningkatan energi awal jauh lebih besar daripada apa yang mencapai blok biaryl. Jadi, hanya 72% darinya yang ditransmisikan dari motor ke blok biaryl.Pengamatan di atas menunjukkan bahwa panjang hubungan antara motor dan blok biaryl merupakan aspek penting dalam proses mengubah tingkat transfer energi. Oleh karena itu, koneksi yang lebih pendek memungkinkan lebih banyak energi potensial ditransfer dari BR dan DR ke AT dan CT.Untuk seorang kenalan yang lebih mendetail dengan nuansa penelitian ini, saya sarankan Anda melihat laporan para ilmuwan dan bahan tambahan untuk itu.Epilog
Dalam karya ini, para ilmuwan menggambarkan prinsip operasi penemuan mereka, berdasarkan interaksi motor biologis dan elemen molekuler. Instalasi yang dikembangkan memungkinkan Anda untuk mentransfer energi potensial dari motor ke elemen tertentu (dalam hal ini, itu adalah blok biaryl). Sebagai hasil dari aktivitas motorik, atropisomerisasi termal biaryl dipaksa untuk melanjutkan tanpa arah dan pada kecepatan yang lebih cepat. Transfer energi terjadi selama ratchet yang diaktifkan secara termal (langkah-langkah) dalam mekanisme rotasi. Juga ditemukan bahwa sekitar 72% energi awal motor ditransmisikan ke unit tertentu.Penemuan ini merupakan langkah penting menuju penciptaan perangkat nanometer biologis lengkap. Penggunaan bio-motor dalam desain mekanisme tersebut adalah karena biokompatibilitasnya, kemudahan perubahan fungsi karena rekayasa genetika, serta skala nano alami mereka. Motor buatan pada saat ini tidak dapat membanggakan kombinasi keuntungan di atas.Para peneliti mengatakan bahwa langkah selanjutnya dalam pekerjaan mereka adalah menunjukkan bahwa instalasi mereka tidak hanya dapat melakukan tindakan abstrak, tetapi juga bermanfaat. Dengan kata lain, kreasi mereka di masa depan akan lebih disempurnakan untuk melakukan fungsi yang bermanfaat di tingkat molekuler.Terima kasih atas perhatian Anda, tetap ingin tahu dan selamat bekerja, kawan. :)Sedikit iklan :)
Terima kasih untuk tetap bersama kami. Apakah Anda suka artikel kami? Ingin melihat materi yang lebih menarik? Dukung kami dengan melakukan pemesanan atau merekomendasikan kepada teman Anda, cloud VPS untuk pengembang mulai $ 4,99 , analog unik dari server entry-level yang diciptakan oleh kami untuk Anda: Seluruh kebenaran tentang VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps mulai dari $ 19 atau cara membagi server? (opsi tersedia dengan RAID1 dan RAID10, hingga 24 core dan hingga 40GB DDR4).Dell R730xd 2 kali lebih murah di pusat data Equinix Tier IV di Amsterdam? Hanya kami yang memiliki 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV dari $ 199 di Belanda!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - mulai dari $ 99! Baca tentang Cara Membangun Infrastruktur Bldg. kelas c menggunakan server Dell R730xd E5-2650 v4 seharga 9.000 euro untuk satu sen?Source: https://habr.com/ru/post/undefined/
All Articles