Synchronous Fuete: Moteurs biologiques en nanotechnologie
 Dans les vastes étendues de notre galaxie, de nombreux secrets sont cachés que les scientifiques du monde entier s'efforcent de trouver et de percer. Cependant, il n'est pas nécessaire que quelque chose soit gros pour être mystérieux. Une preuve éclatante de cela est le monde situé au niveau cellulaire. Bon nombre des formes les plus diverses, la structure, la fonction et le but des cellules remplissent ensemble une tâche commune - maintenir la vie du corps. Si vous exagérez, les cellules, comme les gens, ont des professions: facteurs, transmettant des informations entre les cellules et les tissus; les gardes-frontières qui identifient et combattent les infections; archivistes collectant et stockant des informations, etc. Dans cette incroyable gamme de spécialités, il y a un métier très inhabituel, du moins pour nous, un moteur biologique, qui génère la force mécanique nécessaire au mouvement des cellules.Ces cellules sont particulièrement intéressantes dans le contexte de la nanotechnologie. Auparavant, il y avait un problème dans la mise en œuvre d'un nano-dispositif fonctionnel basé sur des moteurs biologiques - les moteurs doivent être intégrés dans des systèmes plus grands afin que leurs mouvements mécaniques puissent être efficacement connectés à d'autres unités moléculaires. Des scientifiques de l'Université de Munich (Allemagne) ont réussi à se rapprocher de la mise en œuvre de ce concept. Quelles cellules et unités moléculaires spécifiques ont été utilisées pour créer le modèle, comment leur travail a-t-il été contrôlé, quelle était la tâche du système de travail et quels résultats a-t-il montré? Nous trouverons des réponses à ces questions dans le rapport du groupe de recherche. Aller.
Dans les vastes étendues de notre galaxie, de nombreux secrets sont cachés que les scientifiques du monde entier s'efforcent de trouver et de percer. Cependant, il n'est pas nécessaire que quelque chose soit gros pour être mystérieux. Une preuve éclatante de cela est le monde situé au niveau cellulaire. Bon nombre des formes les plus diverses, la structure, la fonction et le but des cellules remplissent ensemble une tâche commune - maintenir la vie du corps. Si vous exagérez, les cellules, comme les gens, ont des professions: facteurs, transmettant des informations entre les cellules et les tissus; les gardes-frontières qui identifient et combattent les infections; archivistes collectant et stockant des informations, etc. Dans cette incroyable gamme de spécialités, il y a un métier très inhabituel, du moins pour nous, un moteur biologique, qui génère la force mécanique nécessaire au mouvement des cellules.Ces cellules sont particulièrement intéressantes dans le contexte de la nanotechnologie. Auparavant, il y avait un problème dans la mise en œuvre d'un nano-dispositif fonctionnel basé sur des moteurs biologiques - les moteurs doivent être intégrés dans des systèmes plus grands afin que leurs mouvements mécaniques puissent être efficacement connectés à d'autres unités moléculaires. Des scientifiques de l'Université de Munich (Allemagne) ont réussi à se rapprocher de la mise en œuvre de ce concept. Quelles cellules et unités moléculaires spécifiques ont été utilisées pour créer le modèle, comment leur travail a-t-il été contrôlé, quelle était la tâche du système de travail et quels résultats a-t-il montré? Nous trouverons des réponses à ces questions dans le rapport du groupe de recherche. Aller.Base d'étude
Comme mentionné ci-dessus, un moteur biologique est une cellule qui génère la force mécanique nécessaire pour réaliser le mouvement des cellules, ainsi que le transport intracellulaire. Ces moteurs comprennent les protéines motrices et les complexes protéiques.Pour mener à bien ses activités non standard, les protéines motrices, comme toute machine, ont besoin de carburant. L'acide adénosine triphosphorique (ATP, C 10 H 16 N 5 O 13 P 3 ) joue son rôle . L'ATP est une source d'énergie universelle pour tous les processus biologiques des systèmes vivants.Les protéines motrices fonctionnent par hydrolyse * ATP, qui permet aux protéines de convertir l'énergie chimique en travail mécanique.L'hydrolyse * est la réaction de décomposition d'échange entre une substance dissoute et un solvant, dans laquelle la décomposition d'une substance (dans ce cas l'ATP) et de l'eau se produit avec la formation de nouveaux composés.
En particulier, les protéines motrices comprennent les myosines, les kinésines et les dynéines. Les myosines représentent environ 40 à 60% de toutes les protéines musculaires, participant au processus de contraction musculaire.Les kinésines, qui sont devenues un mème scientifique il y a plusieurs années, voyagent à travers des microtubules (structures intracellulaires protéiques), participant aux processus de mitose, de méiose et de transport vésiculaire.Démonstration vidéo de la façon dont la kinésine se déplace à travers un microtubule, transformé en mème. (l'original est tiré de La vie intérieure d'une cellule ; la musique superposée sur la vidéo est Stayin 'Alive, Bee Gees, 1977).Les dynéines, comme les kinésines, se déplacent également le long des microtubules du cytosquelette, participant au processus de transfert de cargaison (vésicules, mitochondries, etc.).L'intérêt pour les biomoteurs de la part des nanotechnologues est dû à plusieurs facteurs importants: l'échelle nanométrique, la biocompatibilité et la capacité d'utiliser le génie génétique pour créer des biomoteurs avec des fonctions spécifiées.À l'heure actuelle, il existe un certain nombre de développements basés sur les principes d'action des moteurs biologiques. Cependant, les informations concernant le travail réel qu'un moteur moléculaire unique peut faire ne sont pas encore suffisantes pour une mise en œuvre complète. Un autre casse-tête pour les scientifiques est la question de savoir comment intégrer le bloc de mouvement moléculaire dans une structure plus grande afin que son mouvement dirigé soit effectivement transformé en une augmentation de l'énergie potentielle à des points éloignés de la structure.De toute évidence, il existe de nombreux obstacles, mais cela n'a jamais empêché les scientifiques. Dans l'étude que nous envisageons aujourd'hui, les scientifiques décrivent l'installation moléculaire 1, qui permet le transfert d'énergie potentielle du bloc moteur vers un récepteur distant, accélérant ainsi le mouvement de ce dernier (image n ° 1).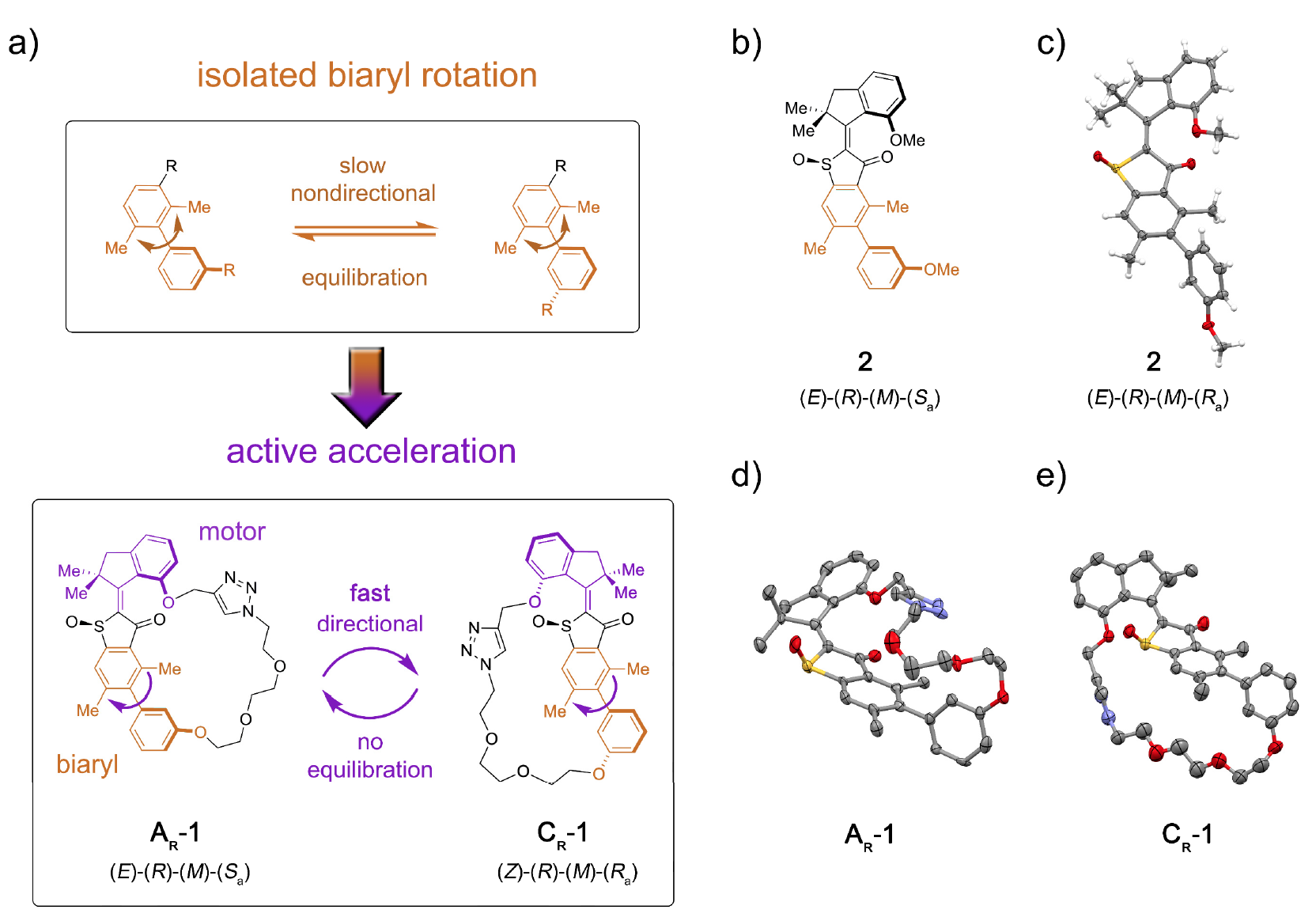 Image n ° 1Le rôle de l'unité réceptrice a été joué par le biaryl * chiral axial qui, dans sa forme non captive (système modèle 2), subit une rotation lente et non directionnelle de l'atropisomérisation dans le sens de l'équilibre des énantiomères 1: 1 ( 1a ).
Image n ° 1Le rôle de l'unité réceptrice a été joué par le biaryl * chiral axial qui, dans sa forme non captive (système modèle 2), subit une rotation lente et non directionnelle de l'atropisomérisation dans le sens de l'équilibre des énantiomères 1: 1 ( 1a ).La chiralité axiale * résulte d'une disposition non plane de substituants par rapport à un axe - l'axe de la chiralité.
Biaryl * - Tout composé contenant une sous-structure, qui est une combinaison de deux composés aromatiques ou groupes aryle, s'ils sont reliés par une seule liaison.
La conjugaison du bloc moteur moléculaire permet unidirectionnellement de détourner cette atropisomérisation de l'équilibre. De ce fait, l'atropisomérisation du biaryl n'est plus passive (suite au fonctionnement du moteur), mais représente un obstacle énergétique contre lequel le moteur doit travailler activement.Dans la machine 1, une augmentation du taux d'atropisomérisation du biaryl de plusieurs ordres de grandeur est obtenue par l'action d'un moteur qui supprime les barrières internes à la rotation isolée du biaryl (système modèle 2 par 1b et 1s ).L'installation moléculaire 1 comprend une unité de moteur moléculaire basée sur HTI, qui appartient à la classe des chromophores indigo. Ce type d'unité est extrêmement sensible à la lumière, c'est-à-dire grâce à la lumière, il peut être contrôlé. Le bloc moteur est connecté de manière covalente à un axe biaryle distant, qui ne se déplace pas avec la rotation du moteur.L'une des différences entre cette installation et ses versions antérieures est la présence d'états supplémentaires (étapes): la version antérieure avait 4 états et n'était régulée que par les étapes de rotation du moteur; une nouvelle option est un système à six vitesses (image n ° 2).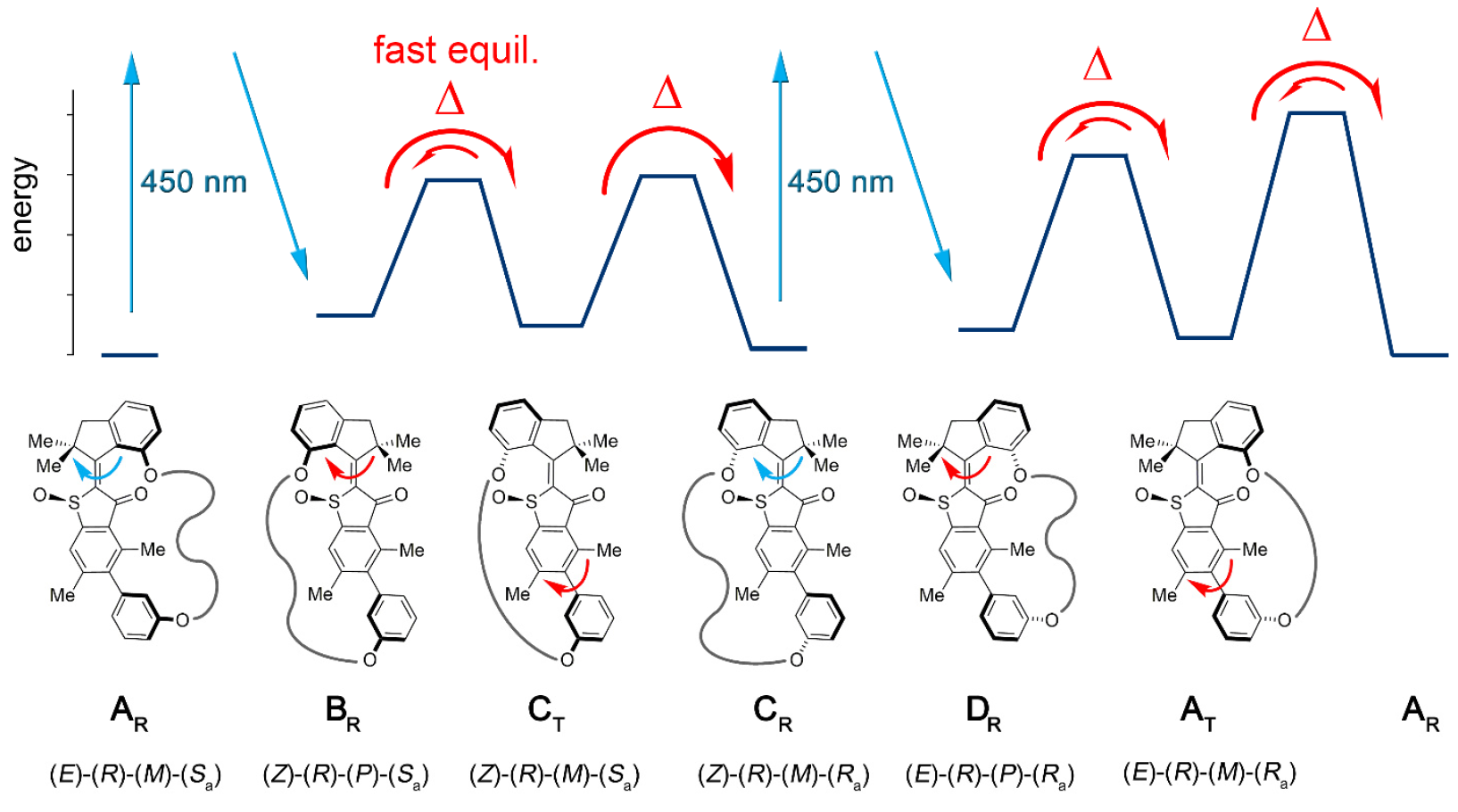 Image n ° 2Six états isomères sont nommés comme suit: AT pour l'état de contrainte A; AR pour l'état détendu A, ainsi que BR, CT, CR et DR (états intermédiaires du système).Cinq de ces étapes peuvent être observées expérimentalement, ce qui confirme la présence de la sixième étape et, par conséquent, confirme la pleine directionnalité pour une rotation de 360 degrés du système associé du moteur de bloc et du récepteur de bloc.Le système moteur 1 a été synthétisé par la méthode convergente en raison du précurseur HTI bromé, auquel une liaison covalente contenant la fonction d'ester boronique a été attachée via une réaction de clic catalysée par le cuivre.Par la suite, la réaction de Suzuki * donne une macrocyclisation * , suivie d'une oxydation pour donner la structure finale 1.
Image n ° 2Six états isomères sont nommés comme suit: AT pour l'état de contrainte A; AR pour l'état détendu A, ainsi que BR, CT, CR et DR (états intermédiaires du système).Cinq de ces étapes peuvent être observées expérimentalement, ce qui confirme la présence de la sixième étape et, par conséquent, confirme la pleine directionnalité pour une rotation de 360 degrés du système associé du moteur de bloc et du récepteur de bloc.Le système moteur 1 a été synthétisé par la méthode convergente en raison du précurseur HTI bromé, auquel une liaison covalente contenant la fonction d'ester boronique a été attachée via une réaction de clic catalysée par le cuivre.Par la suite, la réaction de Suzuki * donne une macrocyclisation * , suivie d'une oxydation pour donner la structure finale 1.La réaction de Suzuki * est une réaction organique des acides aryle et vinylboronique avec des halogénures d'aryle ou de vinyle, catalysée par des complexes Pd (0).
Macrocyclisation * - une réaction de cyclisation qui conduit à la formation d'un macrocycle, c'est-à-dire un composé chimique dans lequel il y a 9 atomes liés ou plus formant un cycle.
Pour les deux états AR et CR les plus stables du système 1 et l'état le plus stable du système 2, des cristaux adaptés à l'analyse structurale ont été obtenus (isomères configurés ® de AR racémique * à 1d ; CR à 1e ).Racemat * est un mélange équimolaire de deux stéréoisomères, qui sont des images miroir l'un de l'autre. Le composé racémique se compose de cristaux, dans chacun desquels se trouvent des molécules des deux énantiomères, et leur rapport est de 1: 1.
Comme AR / AT et CR peuvent être séparés par HPLC (chromatographie liquide à haute performance), leur comportement dans des conditions de chauffage et d'irradiation à différentes températures peut être étudié indépendamment. Lorsque la solution racémique de CR dans (CDCl 2 ) 2 a été chauffée à 80 ° C - 140 ° C, une RA plus stable s'est formée dans 93% des cas. Cela définit la différence d'enthalpie libre ∆G = 1,8–1,9 kcal / mol dans cette plage de température entre ces deux états. L'analyse cinétique correspondante a révélé une barrière à haute énergie de 28,2 kcal / mol à 80 ° C, accompagnant l'isomérisation thermique de la double liaison Z / E.Lors du refroidissement du CR pur à -105 ° C dans CD 2 Cl 2 / CS 2(rapport 4/1) et en l'irradiant avec une lumière de 450 nm, un nouvel ensemble de signaux apparaît, qui diffère de l'ensemble connu de signaux AR et AT ( 3a ). Image n ° 3Les signaux de l'isomère DR se désintègrent à 75% en 28 minutes à une température de -80 ° C et une obscurité totale, mais les signaux de l'isomère AT, au contraire, croissent ( 3 ).L'équilibre thermique entre DR (les 12% restants) et AT (88%) est observé à -60 ° C, ce qui s'exprime en ∆G = 0,84 kcal / mol entre les deux états. L'analyse cinétique de la décomposition thermique a révélé une enthalpie d'activation libre associée ∆ ‡ G = 13,9 kcal / mol à -80 ° C pour ce processus. Aux températures de -40 à 0 ° C, une atténuation des signaux AT et une augmentation concomitante des signaux AR connus sont observées jusqu'à ce que l'équilibre soit atteint entre eux. Une analyse cinétique de ce processus a déterminé ∆ ‡ G égal à 18,4 à 19,3 kcal / mol à des températures de –40 à 0 ° C.Ainsi, il a été constaté que l'irradiation de CR conduit au premier produit de photoisomérisation DR, qui est thermiquement converti en isomère AT en inversant une seule hélice dans le bloc moteur. Lors du refroidissement d'une solution d'équilibre AR / AT dans CD 2Cl 2 / CS 2 jusqu'à -105 ° C et lorsqu'elle a été irradiée avec une lumière de 450 nm, la photoréaction AR a été observée, en raison de sa prévalence. Un nouvel ensemble de signaux apparaît également qui diffère des signaux CR ( 3f ) déjà connus . Ces nouveaux signaux, étant le produit de la photoréaction AR, confirment la direction attendue de la photoisomérisation de AR à BR, qui se stabilise alors encore plus, subissant une inversion hélicoïdale rapide en CT même à basse température.À une température de -80 ° C et dans l'obscurité totale, les signaux BR / CT disparaissent presque complètement et seuls les signaux CR continuent de s'amplifier davantage ( 3e ). Cela donne une limite inférieure pour la différence d'énergie entre CT et CR égale à 0,98 kcal / mol.L'analyse cinétique a déterminé ∆ ‡ G = 13,4–13,5 kcal / mol, ce qui est nécessaire pour la conversion de ST en CR. On peut voir que l'énergie d'activation de l'atropisomérisation du CT en CR diminue par rapport à celle de l'atropisomérisation de l'AT en AR (18,4 kcal / mol). Cela peut être dû à la contrainte (déformation) plus élevée dans le CT par rapport à l'AT, qui peut être due aux grandes distances entre les deux atomes d'oxygène phénoliques, qui servent de points de connexion de la chaîne de liaison dans les structures C, qui ne sont pas dans les structures A. À l'état cristallin, CR entre deux atomes d'oxygène est de 10,6, tandis qu'en AR, il est de 7,6. Cela donne à la chaîne de liaison une plus grande liberté conformationnelle (position spatiale des atomes) dans les structures A que dans les structures C. Si l'on prend en compte l'enthalpie d'activation libre pour la transformation thermique du CT en CR,puis ce processus de conversion peut être complètement arrêté à -105 ° C.À une température de -105 ° C, il est possible de vérifier le degré d'accumulation de CT lors d'une forte exposition à la lumière AR. Cela est possible si la différence d'énergie entre BR et ST dépasse 2,0 kcal / mol et si la structure ST n'est impliquée dans aucun processus de photochimie par elle-même. Cependant, l'accumulation de CT pendant l'irradiation AR à -105 ° C n'est pas possible, car il y a un équilibre thermique rapide entre CT et BR.
Image n ° 3Les signaux de l'isomère DR se désintègrent à 75% en 28 minutes à une température de -80 ° C et une obscurité totale, mais les signaux de l'isomère AT, au contraire, croissent ( 3 ).L'équilibre thermique entre DR (les 12% restants) et AT (88%) est observé à -60 ° C, ce qui s'exprime en ∆G = 0,84 kcal / mol entre les deux états. L'analyse cinétique de la décomposition thermique a révélé une enthalpie d'activation libre associée ∆ ‡ G = 13,9 kcal / mol à -80 ° C pour ce processus. Aux températures de -40 à 0 ° C, une atténuation des signaux AT et une augmentation concomitante des signaux AR connus sont observées jusqu'à ce que l'équilibre soit atteint entre eux. Une analyse cinétique de ce processus a déterminé ∆ ‡ G égal à 18,4 à 19,3 kcal / mol à des températures de –40 à 0 ° C.Ainsi, il a été constaté que l'irradiation de CR conduit au premier produit de photoisomérisation DR, qui est thermiquement converti en isomère AT en inversant une seule hélice dans le bloc moteur. Lors du refroidissement d'une solution d'équilibre AR / AT dans CD 2Cl 2 / CS 2 jusqu'à -105 ° C et lorsqu'elle a été irradiée avec une lumière de 450 nm, la photoréaction AR a été observée, en raison de sa prévalence. Un nouvel ensemble de signaux apparaît également qui diffère des signaux CR ( 3f ) déjà connus . Ces nouveaux signaux, étant le produit de la photoréaction AR, confirment la direction attendue de la photoisomérisation de AR à BR, qui se stabilise alors encore plus, subissant une inversion hélicoïdale rapide en CT même à basse température.À une température de -80 ° C et dans l'obscurité totale, les signaux BR / CT disparaissent presque complètement et seuls les signaux CR continuent de s'amplifier davantage ( 3e ). Cela donne une limite inférieure pour la différence d'énergie entre CT et CR égale à 0,98 kcal / mol.L'analyse cinétique a déterminé ∆ ‡ G = 13,4–13,5 kcal / mol, ce qui est nécessaire pour la conversion de ST en CR. On peut voir que l'énergie d'activation de l'atropisomérisation du CT en CR diminue par rapport à celle de l'atropisomérisation de l'AT en AR (18,4 kcal / mol). Cela peut être dû à la contrainte (déformation) plus élevée dans le CT par rapport à l'AT, qui peut être due aux grandes distances entre les deux atomes d'oxygène phénoliques, qui servent de points de connexion de la chaîne de liaison dans les structures C, qui ne sont pas dans les structures A. À l'état cristallin, CR entre deux atomes d'oxygène est de 10,6, tandis qu'en AR, il est de 7,6. Cela donne à la chaîne de liaison une plus grande liberté conformationnelle (position spatiale des atomes) dans les structures A que dans les structures C. Si l'on prend en compte l'enthalpie d'activation libre pour la transformation thermique du CT en CR,puis ce processus de conversion peut être complètement arrêté à -105 ° C.À une température de -105 ° C, il est possible de vérifier le degré d'accumulation de CT lors d'une forte exposition à la lumière AR. Cela est possible si la différence d'énergie entre BR et ST dépasse 2,0 kcal / mol et si la structure ST n'est impliquée dans aucun processus de photochimie par elle-même. Cependant, l'accumulation de CT pendant l'irradiation AR à -105 ° C n'est pas possible, car il y a un équilibre thermique rapide entre CT et BR. Tableau n ° 1La combinaison des observations et des données ci-dessus vous permet de créer une image mécanique d'une séquence de rotation unidirectionnelle en 6 étapes dans le modèle 1 (image n ° 2 et n ° 4).
Tableau n ° 1La combinaison des observations et des données ci-dessus vous permet de créer une image mécanique d'une séquence de rotation unidirectionnelle en 6 étapes dans le modèle 1 (image n ° 2 et n ° 4). Image n ° 4Des expériences avec différentes conditions de température nous ont permis de déterminer la dépendance en température de ∆G entre CR et AR / AT, DR et AT, AT et AR, ainsi que la dépendance en température de ∆ G pour BR / CT et CR, DR et AT, AT et AT (tableau n ° 1).Contrairement au mécanisme à 4 vitesses habituel basé sur le moteur HTI, la rotation complète à 360 degrés comprend deux étapes supplémentaires, qui sont l'augmentation de l'énergie potentielle transmise du moteur au bloc biaryle. Au cours de ces deux étapes, une liaison énergétique se forme entre le moteur et le bloc biaryle ( 4b ), c'est-à-dire ce dernier n'est plus énergétiquement dégénéré.
Image n ° 4Des expériences avec différentes conditions de température nous ont permis de déterminer la dépendance en température de ∆G entre CR et AR / AT, DR et AT, AT et AR, ainsi que la dépendance en température de ∆ G pour BR / CT et CR, DR et AT, AT et AT (tableau n ° 1).Contrairement au mécanisme à 4 vitesses habituel basé sur le moteur HTI, la rotation complète à 360 degrés comprend deux étapes supplémentaires, qui sont l'augmentation de l'énergie potentielle transmise du moteur au bloc biaryle. Au cours de ces deux étapes, une liaison énergétique se forme entre le moteur et le bloc biaryle ( 4b ), c'est-à-dire ce dernier n'est plus énergétiquement dégénéré. Image n ° 5Au lieu de cela, l'énergie de l'un des atropisomères augmente en raison des étapes de «cliquet» du moteur pendant une rotation complète ( 5 set 5d ). Ce changement d'énergie fournit jusqu'à 90% de l'interconversion de l'atropisomère aux étapes thermiquement activées suivantes et, par conséquent, une rotation presque complète du bloc de biaryle pendant un cycle de fonctionnement du mécanisme.Cette observation peut indiquer que le bloc biaryle a subi une augmentation d'énergie potentielle, qui est le résultat du fonctionnement du bloc moteur. Il convient de noter que l'augmentation initiale de l'énergie est beaucoup plus importante que ce qui atteint le bloc biaryle. Ainsi, seulement 72% de celui-ci est transmis du moteur au bloc biaryle.Les observations ci-dessus suggèrent que la longueur de la connexion entre le moteur et le bloc biaryle est un aspect important dans le processus de changement du degré de transfert d'énergie. Par conséquent, une connexion plus courte peut permettre de transférer plus d'énergie potentielle du BR et du DR vers l'AT et le CT.Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de consulter le rapport des scientifiques et les documents supplémentaires qui s'y rapportent.
Image n ° 5Au lieu de cela, l'énergie de l'un des atropisomères augmente en raison des étapes de «cliquet» du moteur pendant une rotation complète ( 5 set 5d ). Ce changement d'énergie fournit jusqu'à 90% de l'interconversion de l'atropisomère aux étapes thermiquement activées suivantes et, par conséquent, une rotation presque complète du bloc de biaryle pendant un cycle de fonctionnement du mécanisme.Cette observation peut indiquer que le bloc biaryle a subi une augmentation d'énergie potentielle, qui est le résultat du fonctionnement du bloc moteur. Il convient de noter que l'augmentation initiale de l'énergie est beaucoup plus importante que ce qui atteint le bloc biaryle. Ainsi, seulement 72% de celui-ci est transmis du moteur au bloc biaryle.Les observations ci-dessus suggèrent que la longueur de la connexion entre le moteur et le bloc biaryle est un aspect important dans le processus de changement du degré de transfert d'énergie. Par conséquent, une connexion plus courte peut permettre de transférer plus d'énergie potentielle du BR et du DR vers l'AT et le CT.Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de consulter le rapport des scientifiques et les documents supplémentaires qui s'y rapportent.Épilogue
Dans ce travail, les scientifiques ont décrit le principe de fonctionnement de leur invention, basé sur l'interaction des moteurs biologiques et des éléments moléculaires. L'installation développée vous permet de transférer l'énergie potentielle du moteur vers un élément spécifique (dans ce cas, il s'agit d'un bloc biaryle). En raison de l'activité motrice, l'atropisomérisation thermique du biaryl est forcée de se dérouler unidirectionnellement et à un rythme plus rapide. Le transfert d'énergie se produit lors de cliquets activés thermiquement (étapes) dans le mécanisme de rotation. Il a également été constaté qu'environ 72% de l'énergie initiale du moteur est transmise à une unité donnée.Cette invention est une étape importante vers la création de dispositifs nanométriques biologiques complets. L'utilisation de bio-moteurs dans la conception de tels mécanismes est due à leur biocompatibilité, à la facilité de changement de fonctionnalité due au génie génétique, et également à leur nature nanométrique. Les moteurs artificiels ne peuvent actuellement se vanter de la combinaison d'avantages ci-dessus.Les chercheurs disent que la prochaine étape de leur travail sera de démontrer que leur installation est capable d'effectuer non seulement des actions abstraites, mais aussi d'être utile. En d'autres termes, leur future création sera plus affinée pour remplir des fonctions utiles au niveau moléculaire.Merci de votre attention, restez curieux et bonne semaine de travail, les gars. :)Un peu de publicité :)
Merci de rester avec nous. Aimez-vous nos articles? Vous voulez voir des matériaux plus intéressants? Soutenez-nous en passant une commande ou en recommandant à vos amis, le cloud VPS pour les développeurs à partir de 4,99 $ , un analogue unique de serveurs d'entrée de gamme que nous avons inventé pour vous: Toute la vérité sur VPS (KVM) E5-2697 v3 (6 cœurs) 10 Go DDR4 480 Go SSD 1 Gbit / s à partir de 19 $ ou comment diviser le serveur? (les options sont disponibles avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).Dell R730xd 2 fois moins cher au centre de données Equinix Tier IV à Amsterdam? Nous avons seulement 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV à partir de 199 $ aux Pays-Bas!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - à partir de 99 $! En savoir plus sur la création d'un bâtiment d'infrastructure. classe c utilisant des serveurs Dell R730xd E5-2650 v4 coûtant 9 000 euros pour un sou?Source: https://habr.com/ru/post/undefined/
All Articles