Fuete síncrono: motores biológicos en nanotecnología
 En las vastas extensiones de nuestra galaxia, se esconden muchos secretos que los científicos de todo el mundo están tratando de encontrar y desentrañar. Sin embargo, no es necesario que algo sea grande para ser misterioso. Una prueba vívida de esto es el mundo que yace a nivel celular. Muchas de las más diversas en forma, estructura, funcionalidad y propósito de las células juntas cumplen una tarea común: mantener la vida del cuerpo. Si exageras, entonces las células, como las personas, tienen profesiones: carteros, que transmiten información entre células y tejidos; guardias fronterizos que identifican y combaten infecciones; archiveros recolectando y almacenando información, etc. En esta increíble gama de especialidades, existe una profesión muy inusual, al menos para nosotros, un motor biológico, que genera la fuerza mecánica necesaria para el movimiento de las células.Estas células son particularmente interesantes en el contexto de la nanotecnología. Anteriormente, había un problema en la implementación de un nanodispositivo viable basado en motores biológicos: los motores deben integrarse en sistemas más grandes para que sus movimientos mecánicos puedan conectarse efectivamente con otras unidades moleculares. Científicos de la Universidad de Munich (Alemania) lograron acercarse a la implementación de este concepto. ¿Qué células específicas y unidades moleculares se usaron para crear el modelo, cómo se controló su trabajo, cuál fue la tarea del sistema de trabajo y qué resultados mostró? Encontraremos respuestas a estas preguntas en el informe del grupo de investigación. Vamos.
En las vastas extensiones de nuestra galaxia, se esconden muchos secretos que los científicos de todo el mundo están tratando de encontrar y desentrañar. Sin embargo, no es necesario que algo sea grande para ser misterioso. Una prueba vívida de esto es el mundo que yace a nivel celular. Muchas de las más diversas en forma, estructura, funcionalidad y propósito de las células juntas cumplen una tarea común: mantener la vida del cuerpo. Si exageras, entonces las células, como las personas, tienen profesiones: carteros, que transmiten información entre células y tejidos; guardias fronterizos que identifican y combaten infecciones; archiveros recolectando y almacenando información, etc. En esta increíble gama de especialidades, existe una profesión muy inusual, al menos para nosotros, un motor biológico, que genera la fuerza mecánica necesaria para el movimiento de las células.Estas células son particularmente interesantes en el contexto de la nanotecnología. Anteriormente, había un problema en la implementación de un nanodispositivo viable basado en motores biológicos: los motores deben integrarse en sistemas más grandes para que sus movimientos mecánicos puedan conectarse efectivamente con otras unidades moleculares. Científicos de la Universidad de Munich (Alemania) lograron acercarse a la implementación de este concepto. ¿Qué células específicas y unidades moleculares se usaron para crear el modelo, cómo se controló su trabajo, cuál fue la tarea del sistema de trabajo y qué resultados mostró? Encontraremos respuestas a estas preguntas en el informe del grupo de investigación. Vamos.Base de estudio
Como se mencionó anteriormente, un motor biológico es una célula que genera la fuerza mecánica necesaria para realizar el movimiento de las células, así como el transporte intracelular. Estos motores incluyen proteínas motoras y complejos de proteínas.Para llevar a cabo sus actividades no estándar, las proteínas motoras, como cualquier máquina, necesitan combustible. El ácido trifosfórico de adenosina (ATP, C 10 H 16 N 5 O 13 P 3 ) desempeña su papel . El ATP es una fuente universal de energía para todos los procesos biológicos dentro de los sistemas vivos.Las proteínas motoras funcionan por hidrólisis * ATP, que permite a las proteínas convertir la energía química en trabajo mecánico.La hidrólisis * es la reacción de descomposición de intercambio entre una sustancia disuelta y un solvente, en la cual la descomposición de una sustancia (en este caso ATP) y el agua ocurre con la formación de nuevos compuestos.
En particular, las proteínas motoras incluyen miosinas, quinesinas y dineínas. Las miosinas constituyen aproximadamente el 40-60% de todas las proteínas musculares, participando en el proceso de contracción muscular.Las kinesinas, que se convirtieron en un meme científico hace varios años, viajan a través de microtúbulos (estructuras intracelulares de proteínas), participando en los procesos de mitosis, meiosis y transporte vesicular.Video de demostración de cómo la kinesina se mueve a través de un microtúbulo, transformada en un meme. (el original está tomado de The Inner Life of a Cell ; la música superpuesta en el video es Stayin 'Alive, Bee Gees, 1977).Las dineínas, como las kinesinas, también se mueven a lo largo de los microtúbulos del citoesqueleto, participando en el proceso de transferencia de carga (vesículas, mitocondrias, etc.).El interés en los biomotores por parte de los nanotecnólogos se debe a varios factores importantes: nanoescala, biocompatibilidad y la capacidad de utilizar la ingeniería genética para crear biomotores con funciones específicas.En este momento, hay una serie de desarrollos basados en los principios de acción de los motores biológicos. Sin embargo, la información sobre cuánto trabajo real puede hacer un solo motor molecular aún no es suficiente para una implementación completa. Otro enigma para los científicos es la cuestión de cómo integrar el bloque de movimiento molecular en una estructura más grande para que su movimiento dirigido se transforme efectivamente en un aumento de energía potencial en puntos remotos de la estructura.Obviamente, hay muchos obstáculos, pero esto nunca ha detenido a los científicos. En el estudio que estamos considerando hoy, los científicos describen la instalación molecular 1, que permite la transferencia de energía potencial de la unidad motora a una unidad receptora remota, acelerando así el movimiento de esta última (imagen No. 1).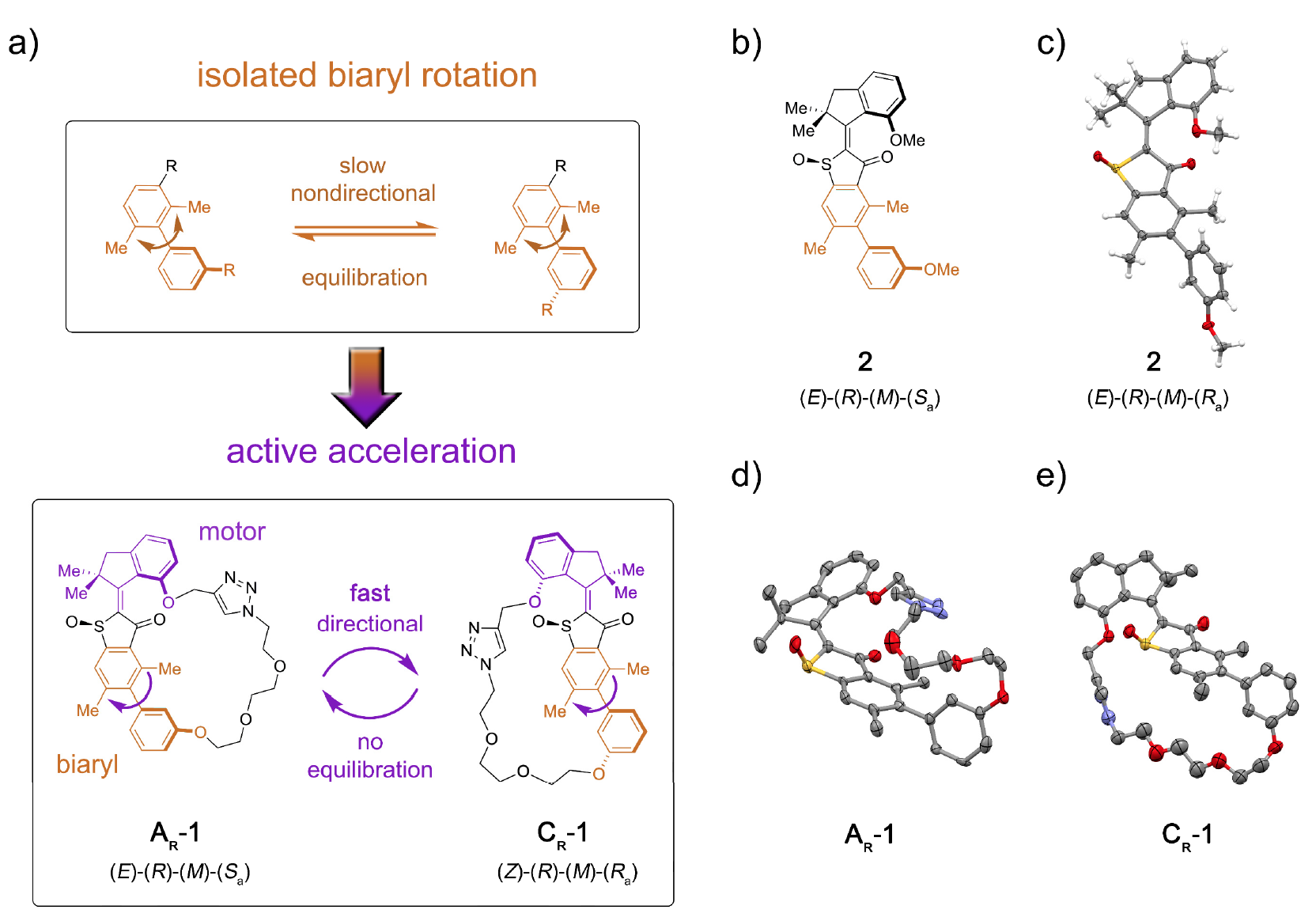 Imagen No. 1El papel de la unidad receptora fue desempeñado por axial-quiral * biarilo * , que en su forma no atada (sistema modelo 2) sufre una rotación lenta y no direccional de atropisomerización en la dirección del equilibrio de los enantiómeros 1: 1 ( 1a ).
Imagen No. 1El papel de la unidad receptora fue desempeñado por axial-quiral * biarilo * , que en su forma no atada (sistema modelo 2) sufre una rotación lenta y no direccional de atropisomerización en la dirección del equilibrio de los enantiómeros 1: 1 ( 1a ).La quiralidad axial * surge como resultado de una disposición no plana de sustituyentes en relación con algún eje, el eje de la quiralidad.
Biarilo * : cualquier compuesto que contenga una subestructura, que es una combinación de dos compuestos aromáticos o grupos arilo, si están conectados por un enlace sencillo.
La conjugación del bloque motor molecular permite que unidireccional desvíe esta atropisomerización del equilibrio. Como resultado, la atropisomerización del biarilo ya no es pasiva (después del funcionamiento del motor), sino que representa un obstáculo energético contra el cual el motor debe trabajar activamente.En la máquina 1, un aumento en la tasa de atropisomerización de biarilo en varios órdenes de magnitud se logra mediante la acción de un motor que suprime las barreras internas a la rotación aislada de biarilo (sistema modelo 2 por 1b y 1s ).La instalación molecular 1 incluye una unidad motora molecular basada en HTI, que pertenece a la clase de cromóforos índigo. Este tipo de unidad es extremadamente sensible a la luz, es decir Gracias a la luz, se puede controlar. El bloque motor está conectado covalentemente a un eje biarilo remoto, que no se mueve con la rotación del motor.Una de las diferencias entre esta instalación y sus versiones anteriores es la presencia de estados adicionales (pasos): la versión anterior tenía 4 estados y solo estaba regulada por los pasos de rotación del motor; Una nueva opción es un sistema de seis velocidades (imagen No. 2).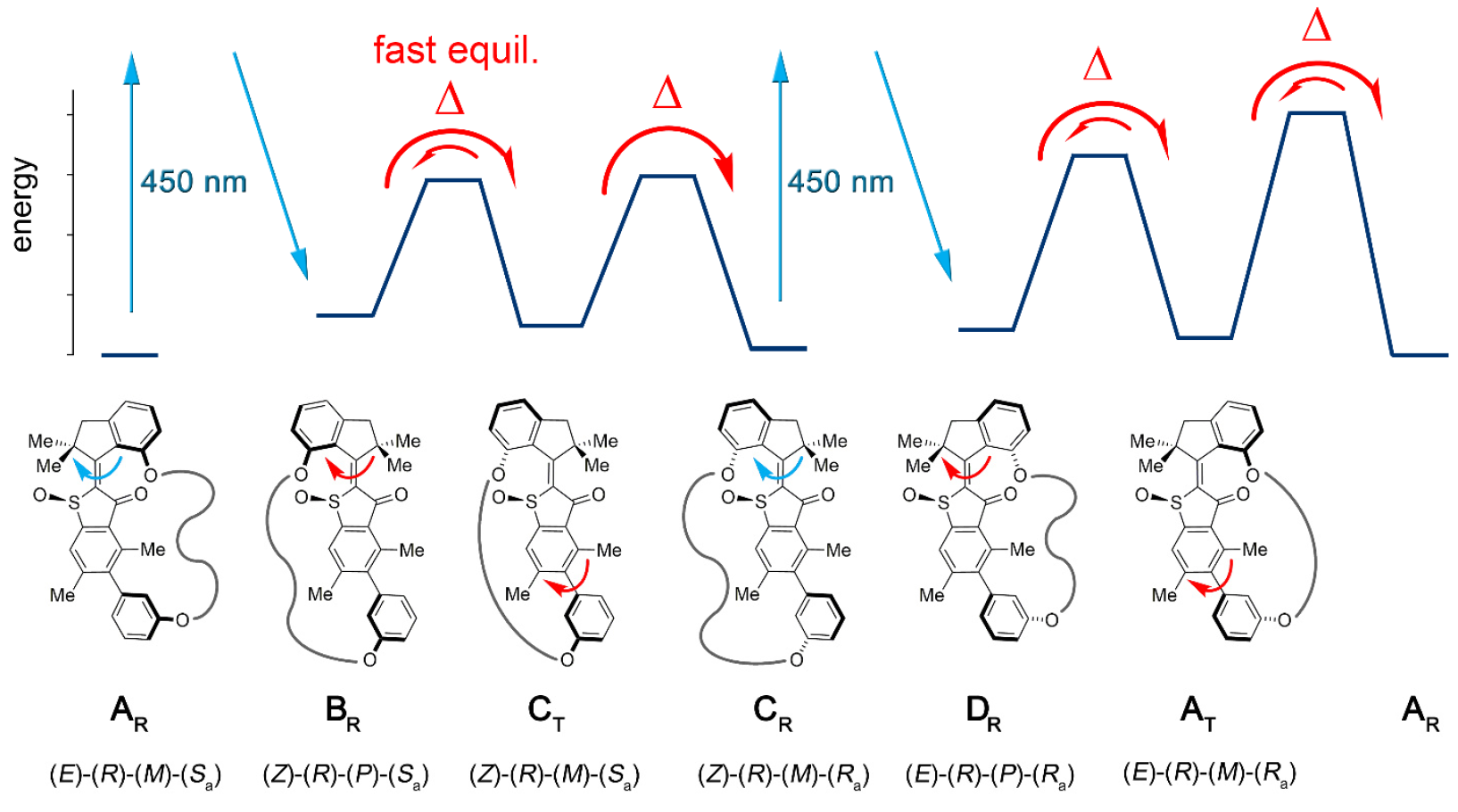 Imagen No. 2Seis estados isoméricos se nombran de la siguiente manera: AT para el estado de estrés A; AR para el estado relajado A, así como BR, CT, CR y DR (estados intermedios del sistema).Se pueden observar cinco de estos pasos experimentalmente, lo que confirma la presencia del sexto paso y, por lo tanto, confirma la direccionalidad completa para una rotación de 360 grados del sistema asociado del motor de bloque y el receptor de bloque.El sistema motor 1 se sintetizó por el método convergente debido al precursor de HTI bromado, al que se unió un enlace covalente que contenía la función del éster borónico a través de una reacción de clic catalizada por cobre.Posteriormente, la reacción de Suzuki * da macrociclación * , seguida de oxidación para dar la estructura final 1.
Imagen No. 2Seis estados isoméricos se nombran de la siguiente manera: AT para el estado de estrés A; AR para el estado relajado A, así como BR, CT, CR y DR (estados intermedios del sistema).Se pueden observar cinco de estos pasos experimentalmente, lo que confirma la presencia del sexto paso y, por lo tanto, confirma la direccionalidad completa para una rotación de 360 grados del sistema asociado del motor de bloque y el receptor de bloque.El sistema motor 1 se sintetizó por el método convergente debido al precursor de HTI bromado, al que se unió un enlace covalente que contenía la función del éster borónico a través de una reacción de clic catalizada por cobre.Posteriormente, la reacción de Suzuki * da macrociclación * , seguida de oxidación para dar la estructura final 1.La reacción de Suzuki * es una reacción orgánica de ácidos aril y vinilborónico con haluros de arilo o vinilo, catalizada por complejos de Pd (0).
Macrociclización * : una reacción de ciclación que conduce a la formación de un macrociclo, es decir Un compuesto químico en el que hay 9 o más átomos unidos formando un anillo.
Para los dos estados más estables de AR y CR del sistema 1 y el estado más estable del sistema 2, se obtuvieron cristales adecuados para el análisis estructural (isómeros de AR racémico * AR configurados en 1d ; CR en 1e ).Racemat * es una mezcla equimolar de dos estereoisómeros, que son imágenes especulares entre sí. El compuesto racémico consiste en cristales, en cada uno de los cuales hay moléculas de ambos enantiómeros, y su proporción es 1: 1.
Dado que AR / AT y CR pueden separarse mediante HPLC (cromatografía líquida de alta resolución), su comportamiento en condiciones de calentamiento e irradiación a diferentes temperaturas puede estudiarse de forma independiente. Cuando la solución de CR racémica en (CDCl 2 ) 2 se calentó a 80 ° C - 140 ° C, se formó un AR más estable en el 93% de los casos. Esto establece la diferencia en la entalpía libre ∆G = 1.8–1.9 kcal / mol en este rango de temperatura entre estos dos estados. El análisis cinético correspondiente reveló una barrera de alta energía de 28.2 kcal / mol a 80 ° C, que acompaña a la isomerización térmica del doble enlace Z / E.Al enfriar CR puro a -105 ° C en CD 2 Cl 2 / CS 2(relación 4/1) e irradiándola con luz de 450 nm, aparece un nuevo conjunto de señales, que difiere del conjunto conocido de señales AR y AT ( 3a ). Imagen n. ° 3Las señales del isómero DR disminuyen al 75% en 28 minutos a una temperatura de -80 ° C y completa oscuridad, pero las señales del isómero AT aumentan, por el contrario ( 3 ).El equilibrio térmico entre DR (el 12% restante) y AT (88%) se observa a -60 ° C, que se expresa en ∆G = 0,84 kcal / mol entre los dos estados. El análisis cinético de la descomposición térmica reveló una entalpía de activación libre acompañante ∆ ‡ G = 13.9 kcal / mol a -80 ° C para este proceso. A temperaturas de -40 a 0 ° C, se observa la atenuación de las señales AT y un aumento concomitante de las señales AR conocidas hasta que se alcanza el equilibrio entre ellas. Un análisis cinético de este proceso determinó ∆ ‡ G igual a 18.4 a 19.3 kcal / mol a temperaturas de –40 ... 0 ° C.Por lo tanto, se descubrió que la irradiación de CR conduce al primer producto de fotoisomerización DR, que se convierte térmicamente en el isómero AT al invertir una hélice única en el bloque motor. Al enfriar una solución de equilibrio de AR / AT en CD 2Cl 2 / CS 2 hasta -105 ° C y cuando se irradió con luz de 450 nm, se observó la fotorreacción AR, debido a su prevalencia. También aparece un nuevo conjunto de señales que difieren de las señales CR ( 3f ) ya conocidas . Estas nuevas señales, siendo el producto de la fotorreacción AR, confirman la dirección esperada de la fotoisomerización de AR a BR, que luego se estabiliza aún más, experimentando una rápida inversión helicoidal a CT incluso a baja temperatura.A una temperatura de -80 ° C y en completa oscuridad, las señales BR / CT desaparecen casi por completo, y solo las señales CR continúan amplificándose aún más ( 3e ). Esto proporciona un límite inferior para la diferencia de energía entre CT y CR igual a 0,98 kcal / mol.El análisis cinético determinó ∆ ‡ G = 13.4–13.5 kcal / mol, que es necesario para la conversión de ST en CR. Se ve que la energía de activación de la atropisomerización de CT en CR disminuye en comparación con la de la atropisomerización de AT en AR (18.4 kcal / mol). Esto puede deberse a la mayor tensión (tensión) en la TC en comparación con la AT, que puede deberse a las grandes distancias entre los dos átomos de oxígeno fenólico, que sirven como puntos de conexión de la cadena de enlace en las estructuras C, que no está en las estructuras A. En el estado cristalino, CR entre dos átomos de oxígeno es 10.6, mientras que en AR es 7.6. Esto le da a la cadena de enlace mayor libertad conformacional (posición espacial de los átomos) en las estructuras A que en las estructuras C. Si tenemos en cuenta la entalpía de activación libre para la transformación térmica de CT en CR,entonces este proceso de conversión se puede detener por completo a -105 ° C.A una temperatura de -105 ° C, es posible verificar el grado de acumulación de CT durante una fuerte exposición a la luz AR. Esto es posible si la diferencia de energía entre BR y ST excede 2.0 kcal / mol y si la estructura ST no está involucrada en ningún proceso de fotoquímica por sí misma. Sin embargo, la acumulación de CT durante la irradiación de AR a -105 ° C no es posible, ya que existe un rápido equilibrio térmico entre CT y BR.
Imagen n. ° 3Las señales del isómero DR disminuyen al 75% en 28 minutos a una temperatura de -80 ° C y completa oscuridad, pero las señales del isómero AT aumentan, por el contrario ( 3 ).El equilibrio térmico entre DR (el 12% restante) y AT (88%) se observa a -60 ° C, que se expresa en ∆G = 0,84 kcal / mol entre los dos estados. El análisis cinético de la descomposición térmica reveló una entalpía de activación libre acompañante ∆ ‡ G = 13.9 kcal / mol a -80 ° C para este proceso. A temperaturas de -40 a 0 ° C, se observa la atenuación de las señales AT y un aumento concomitante de las señales AR conocidas hasta que se alcanza el equilibrio entre ellas. Un análisis cinético de este proceso determinó ∆ ‡ G igual a 18.4 a 19.3 kcal / mol a temperaturas de –40 ... 0 ° C.Por lo tanto, se descubrió que la irradiación de CR conduce al primer producto de fotoisomerización DR, que se convierte térmicamente en el isómero AT al invertir una hélice única en el bloque motor. Al enfriar una solución de equilibrio de AR / AT en CD 2Cl 2 / CS 2 hasta -105 ° C y cuando se irradió con luz de 450 nm, se observó la fotorreacción AR, debido a su prevalencia. También aparece un nuevo conjunto de señales que difieren de las señales CR ( 3f ) ya conocidas . Estas nuevas señales, siendo el producto de la fotorreacción AR, confirman la dirección esperada de la fotoisomerización de AR a BR, que luego se estabiliza aún más, experimentando una rápida inversión helicoidal a CT incluso a baja temperatura.A una temperatura de -80 ° C y en completa oscuridad, las señales BR / CT desaparecen casi por completo, y solo las señales CR continúan amplificándose aún más ( 3e ). Esto proporciona un límite inferior para la diferencia de energía entre CT y CR igual a 0,98 kcal / mol.El análisis cinético determinó ∆ ‡ G = 13.4–13.5 kcal / mol, que es necesario para la conversión de ST en CR. Se ve que la energía de activación de la atropisomerización de CT en CR disminuye en comparación con la de la atropisomerización de AT en AR (18.4 kcal / mol). Esto puede deberse a la mayor tensión (tensión) en la TC en comparación con la AT, que puede deberse a las grandes distancias entre los dos átomos de oxígeno fenólico, que sirven como puntos de conexión de la cadena de enlace en las estructuras C, que no está en las estructuras A. En el estado cristalino, CR entre dos átomos de oxígeno es 10.6, mientras que en AR es 7.6. Esto le da a la cadena de enlace mayor libertad conformacional (posición espacial de los átomos) en las estructuras A que en las estructuras C. Si tenemos en cuenta la entalpía de activación libre para la transformación térmica de CT en CR,entonces este proceso de conversión se puede detener por completo a -105 ° C.A una temperatura de -105 ° C, es posible verificar el grado de acumulación de CT durante una fuerte exposición a la luz AR. Esto es posible si la diferencia de energía entre BR y ST excede 2.0 kcal / mol y si la estructura ST no está involucrada en ningún proceso de fotoquímica por sí misma. Sin embargo, la acumulación de CT durante la irradiación de AR a -105 ° C no es posible, ya que existe un rápido equilibrio térmico entre CT y BR. Tabla No. 1La combinación de las observaciones y datos anteriores le permite crear una imagen mecánica de una secuencia de 6 pasos de rotación unidireccional en el modelo 1 (imagen No. 2 y No. 4).
Tabla No. 1La combinación de las observaciones y datos anteriores le permite crear una imagen mecánica de una secuencia de 6 pasos de rotación unidireccional en el modelo 1 (imagen No. 2 y No. 4). Imagen No. 4Los experimentos con diferentes condiciones de temperatura nos permitieron determinar la dependencia de la temperatura de ∆G entre CR y AR / AT, DR y AT, AT y AR, así como la dependencia de la temperatura de ∆ G para BR / CT y CR, DR y AT, AT y AR (tabla no. 1)A diferencia del mecanismo habitual de 4 velocidades basado en el motor HTI, la rotación completa de 360 grados incluye dos pasos adicionales, que son el aumento de la energía potencial transmitida desde el motor al bloque biarilo. Durante estos dos pasos, se forma un enlace de energía entre el motor y el bloque de biarilo ( 4b ), es decir, este último ya no es degenerado energéticamente.
Imagen No. 4Los experimentos con diferentes condiciones de temperatura nos permitieron determinar la dependencia de la temperatura de ∆G entre CR y AR / AT, DR y AT, AT y AR, así como la dependencia de la temperatura de ∆ G para BR / CT y CR, DR y AT, AT y AR (tabla no. 1)A diferencia del mecanismo habitual de 4 velocidades basado en el motor HTI, la rotación completa de 360 grados incluye dos pasos adicionales, que son el aumento de la energía potencial transmitida desde el motor al bloque biarilo. Durante estos dos pasos, se forma un enlace de energía entre el motor y el bloque de biarilo ( 4b ), es decir, este último ya no es degenerado energéticamente. Imagen No. 5En cambio, la energía de cualquiera de los atropisómeros aumenta debido a los pasos de "trinquete" del motor durante una rotación completa ( 5sy 5d ). Este cambio de energía proporciona hasta el 90% de la interconversión del atropisómero en las siguientes etapas activadas térmicamente y, por lo tanto, una rotación casi completa del bloqueo de biarilo durante un ciclo de trabajo del mecanismo.Esta observación puede indicar que el bloque biarilo ha experimentado un aumento en la energía potencial, que es el resultado del funcionamiento del bloque motor. Vale la pena señalar que el aumento inicial de energía es mucho mayor que el que alcanza el bloque de biarilo. Entonces, solo el 72% se transmite del motor al bloque biarilo.Las observaciones anteriores sugieren que la longitud de la conexión entre el motor y el bloque biarilo es un aspecto importante en el proceso de cambiar el grado de transferencia de energía. Por lo tanto, una conexión más corta puede permitir que se transfiera más energía potencial de BR y DR a AT y CT.Para un conocimiento más detallado de los matices del estudio, le recomiendo que consulte el informe de los científicos y materiales adicionales .
Imagen No. 5En cambio, la energía de cualquiera de los atropisómeros aumenta debido a los pasos de "trinquete" del motor durante una rotación completa ( 5sy 5d ). Este cambio de energía proporciona hasta el 90% de la interconversión del atropisómero en las siguientes etapas activadas térmicamente y, por lo tanto, una rotación casi completa del bloqueo de biarilo durante un ciclo de trabajo del mecanismo.Esta observación puede indicar que el bloque biarilo ha experimentado un aumento en la energía potencial, que es el resultado del funcionamiento del bloque motor. Vale la pena señalar que el aumento inicial de energía es mucho mayor que el que alcanza el bloque de biarilo. Entonces, solo el 72% se transmite del motor al bloque biarilo.Las observaciones anteriores sugieren que la longitud de la conexión entre el motor y el bloque biarilo es un aspecto importante en el proceso de cambiar el grado de transferencia de energía. Por lo tanto, una conexión más corta puede permitir que se transfiera más energía potencial de BR y DR a AT y CT.Para un conocimiento más detallado de los matices del estudio, le recomiendo que consulte el informe de los científicos y materiales adicionales .Epílogo
En este trabajo, los científicos describieron el principio de funcionamiento de su invención, basado en la interacción de motores biológicos y elementos moleculares. La instalación desarrollada le permite transferir energía potencial del motor a un elemento específico (en este caso, es un bloque de biarilo). Como resultado de la actividad motora, la atropisomerización térmica del biarilo se ve obligada a proceder de forma unidireccional y a un ritmo más rápido. La transferencia de energía ocurre durante los trinquetes (pasos) activados térmicamente en el mecanismo de rotación. También se descubrió que aproximadamente el 72% de la energía inicial del motor se transmite a una unidad determinada.Esta invención es un paso importante hacia la creación de dispositivos nanómetros biológicos completos. El uso de motores biológicos en el diseño de tales mecanismos se debe a su biocompatibilidad, facilidad de cambio en la funcionalidad debido a la ingeniería genética, así como a su nanoescala natural. Los motores artificiales en este momento no pueden presumir de la combinación de ventajas anterior.Los investigadores dicen que el siguiente paso en su trabajo será demostrar que su instalación puede realizar no solo acciones abstractas, sino también ser útil. En otras palabras, su futura creación será más refinada para realizar funciones útiles a nivel molecular.Gracias por su atención, sigan curiosos y tengan una buena semana laboral, muchachos. :)Un poco de publicidad :)
Gracias por estar con nosotros. ¿Te gustan nuestros artículos? ¿Quieres ver más materiales interesantes? Apóyenos haciendo un pedido o recomendando a sus amigos VPS basado en la nube para desarrolladores desde $ 4.99 , un análogo único de servidores de nivel básico que inventamos para usted: toda la verdad sobre VPS (KVM) E5-2697 v3 (6 núcleos) 10GB DDR4 480GB SSD 1Gbps desde $ 19 o cómo dividir el servidor? (las opciones están disponibles con RAID1 y RAID10, hasta 24 núcleos y hasta 40GB DDR4).Dell R730xd 2 veces más barato en el centro de datos Equinix Tier IV en Amsterdam? ¡Solo tenemos 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV desde $ 199 en los Países Bajos!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - ¡desde $ 99! Lea sobre Cómo construir un edificio de infraestructura. clase c con servidores Dell R730xd E5-2650 v4 que cuestan 9,000 euros por un centavo?Source: https://habr.com/ru/post/undefined/
All Articles