Synchroner Fuete: Biologische Motoren in der Nanotechnologie
 In den Weiten unserer Galaxie sind viele Geheimnisse verborgen, die Wissenschaftler aus aller Welt so sehr versuchen, zu finden und zu enträtseln. Es ist jedoch nicht notwendig, dass etwas groß ist, um mysteriös zu sein. Ein anschaulicher Beweis dafür ist die Welt, die auf zellulärer Ebene liegt. Viele der verschiedensten in Form, Struktur, Funktion und Zweck der Zellen erfüllen gemeinsam eine gemeinsame Aufgabe - die Erhaltung des Körperlebens. Wenn Sie übertreiben, haben die Zellen wie Menschen Berufe: Postboten, die Informationen zwischen Zellen und Geweben übertragen; Grenzschutzbeamte, die Infektionen identifizieren und bekämpfen; Archivare sammeln und speichern Informationen usw. In diesem unglaublichen Spektrum an Spezialitäten gibt es zumindest für uns einen sehr ungewöhnlichen Beruf - einen biologischen Motor, der die für die Bewegung von Zellen notwendige mechanische Kraft erzeugt.Diese Zellen sind im Kontext der Nanotechnologie besonders interessant. Bisher gab es ein Problem bei der Implementierung eines funktionsfähigen Nanogeräts auf der Basis biologischer Motoren - Motoren müssen in größere Systeme integriert werden, damit ihre mechanischen Bewegungen effektiv mit anderen molekularen Einheiten verbunden werden können. Wissenschaftlern der Universität München gelang es, der Umsetzung dieses Konzepts näher zu kommen. Welche spezifischen Zellen und molekularen Einheiten wurden bei der Erstellung des Modells verwendet, wie wurde ihre Arbeit gesteuert, was war die Aufgabe des Arbeitssystems und welche Ergebnisse zeigte es? Antworten auf diese Fragen finden wir im Bericht der Forschungsgruppe. Gehen.
In den Weiten unserer Galaxie sind viele Geheimnisse verborgen, die Wissenschaftler aus aller Welt so sehr versuchen, zu finden und zu enträtseln. Es ist jedoch nicht notwendig, dass etwas groß ist, um mysteriös zu sein. Ein anschaulicher Beweis dafür ist die Welt, die auf zellulärer Ebene liegt. Viele der verschiedensten in Form, Struktur, Funktion und Zweck der Zellen erfüllen gemeinsam eine gemeinsame Aufgabe - die Erhaltung des Körperlebens. Wenn Sie übertreiben, haben die Zellen wie Menschen Berufe: Postboten, die Informationen zwischen Zellen und Geweben übertragen; Grenzschutzbeamte, die Infektionen identifizieren und bekämpfen; Archivare sammeln und speichern Informationen usw. In diesem unglaublichen Spektrum an Spezialitäten gibt es zumindest für uns einen sehr ungewöhnlichen Beruf - einen biologischen Motor, der die für die Bewegung von Zellen notwendige mechanische Kraft erzeugt.Diese Zellen sind im Kontext der Nanotechnologie besonders interessant. Bisher gab es ein Problem bei der Implementierung eines funktionsfähigen Nanogeräts auf der Basis biologischer Motoren - Motoren müssen in größere Systeme integriert werden, damit ihre mechanischen Bewegungen effektiv mit anderen molekularen Einheiten verbunden werden können. Wissenschaftlern der Universität München gelang es, der Umsetzung dieses Konzepts näher zu kommen. Welche spezifischen Zellen und molekularen Einheiten wurden bei der Erstellung des Modells verwendet, wie wurde ihre Arbeit gesteuert, was war die Aufgabe des Arbeitssystems und welche Ergebnisse zeigte es? Antworten auf diese Fragen finden wir im Bericht der Forschungsgruppe. Gehen.Studienbasis
Wie oben erwähnt, ist ein biologischer Motor eine Zelle, die die mechanische Kraft erzeugt, die erforderlich ist, um die Bewegung von Zellen sowie den intrazellulären Transport zu realisieren. Diese Motoren umfassen Motorproteine und Proteinkomplexe.Motorproteine benötigen wie jede Maschine Kraftstoff, um ihre nicht standardmäßigen Aktivitäten auszuführen. Adenosintriphosphorsäure (ATP, C 10 H 16 N 5 O 13 P 3 ) spielt seine Rolle . ATP ist eine universelle Energiequelle für alle biologischen Prozesse in lebenden Systemen.Motorproteine funktionieren durch Hydrolyse * ATP, wodurch Proteine chemische Energie in mechanische Arbeit umwandeln können.Hydrolyse * ist die Reaktion der Austauschzersetzung zwischen einer gelösten Substanz und einem Lösungsmittel, bei der die Zersetzung einer Substanz (in diesem Fall ATP) und von Wasser unter Bildung neuer Verbindungen erfolgt.
Zu den Motorproteinen gehören insbesondere Myosine, Kinesine und Dyneine. Myosine machen etwa 40-60% aller Muskelproteine aus und sind am Prozess der Muskelkontraktion beteiligt.Kinesine, die vor einigen Jahren zu einem wissenschaftlichen Mem wurden, wandern durch Mikrotubuli (intrazelluläre Proteinstrukturen) und sind an den Prozessen Mitose, Meiose und vesikulärem Transport beteiligt.Videodemonstration, wie sich Kinesin durch ein Mikrotubulus bewegt, das in ein Mem umgewandelt wurde. (Das Original stammt aus dem inneren Leben einer Zelle ; die Musik, die dem Video überlagert ist, ist Stayin 'Alive, Bee Gees, 1977).Dyneins bewegen sich wie Kinesine auch entlang der Mikrotubuli des Zytoskeletts und sind am Prozess des Frachttransfers beteiligt (Vesikel, Mitochondrien usw.).Das Interesse von Nanotechnologen an Biomotoren beruht auf mehreren wichtigen Faktoren: Nanoskala, Biokompatibilität und die Fähigkeit, mithilfe der Gentechnik Biomotoren mit bestimmten Funktionen herzustellen.Derzeit gibt es eine Reihe von Entwicklungen, die auf den Wirkprinzipien biologischer Motoren beruhen. Informationen darüber, wie viel reale Arbeit ein einzelner molekularer Motor leisten kann, reichen jedoch noch nicht für eine vollständige Implementierung aus. Ein weiteres Rätsel für Wissenschaftler ist die Frage, wie der molekulare Bewegungsblock in eine größere Struktur integriert werden kann, damit seine gerichtete Bewegung an entfernten Punkten in der Struktur effektiv in eine Erhöhung der potentiellen Energie umgewandelt wird.Natürlich gibt es viele Hindernisse, aber dies hat Wissenschaftler nie aufgehalten. In der Studie, die wir heute untersuchen, beschreiben Wissenschaftler die molekulare Installation 1, mit der Sie potenzielle Energie von der Motoreinheit auf eine entfernte Empfängereinheit übertragen und so deren Bewegung beschleunigen können (Bild Nr. 1).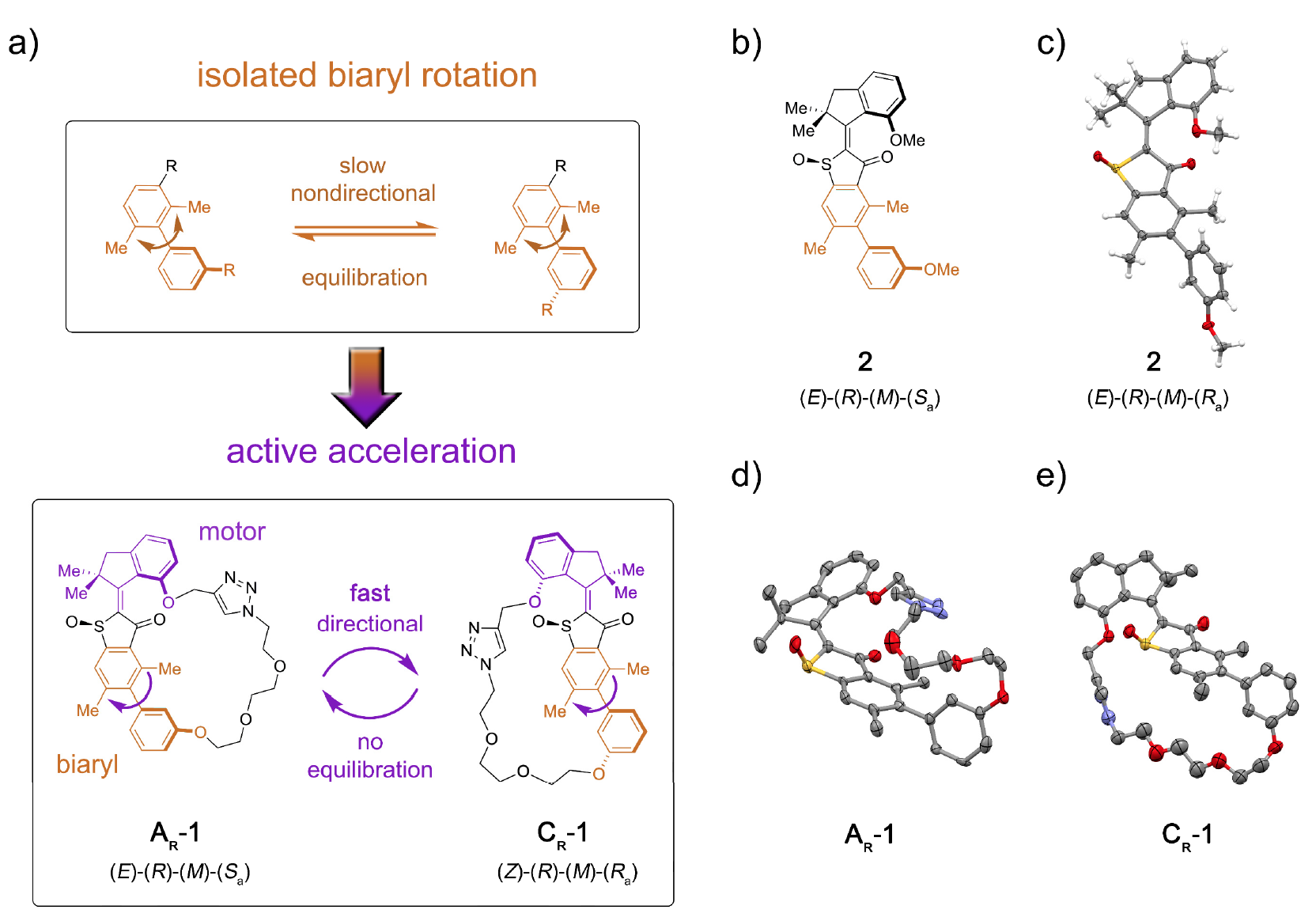 Bild Nr. 1Die Rolle der Empfängereinheit spielte axial-chirales * Biaryl * , das in seiner nicht gebundenen Form (Modellsystem 2) eine langsame und ungerichtete Rotation der Atropisomerisierung in Richtung des Gleichgewichts der Enantiomere 1: 1 ( 1a ) erfährt .
Bild Nr. 1Die Rolle der Empfängereinheit spielte axial-chirales * Biaryl * , das in seiner nicht gebundenen Form (Modellsystem 2) eine langsame und ungerichtete Rotation der Atropisomerisierung in Richtung des Gleichgewichts der Enantiomere 1: 1 ( 1a ) erfährt .Axiale Chiralität * entsteht durch eine nichtplanare Anordnung von Substituenten relativ zu einer Achse - der Achse der Chiralität.
Biaryl * - Jede Verbindung, die eine Unterstruktur enthält, die eine Kombination aus zwei aromatischen Verbindungen oder Arylgruppen ist, wenn sie durch eine Einfachbindung verbunden sind.
Die Konjugation des molekularen Motorblocks ermöglicht es unidirektional, diese Atropisomerisierung vom Gleichgewicht abzulenken. Infolgedessen ist die Atropisomerisierung von Biaryl nicht länger passiv (nach dem Betrieb des Motors), sondern stellt ein Energiehindernis dar, gegen das der Motor aktiv arbeiten muss.In Maschine 1 wird eine Erhöhung der Atropisomerisierungsrate von Biaryl um mehrere Größenordnungen durch die Wirkung eines Motors erreicht, der interne Barrieren für die isolierte Rotation von Biaryl unterdrückt (Modellsystem 2 um 1b und 1s ).Die molekulare Installation 1 enthält eine molekulare Motoreinheit auf Basis von HTI, die zur Klasse der Indigo-Chromophore gehört. Diese Art von Einheit ist extrem lichtempfindlich, d.h. Dank des Lichts kann es gesteuert werden. Der Motorblock ist kovalent mit einer entfernten Biarylachse verbunden, die sich nicht mit der Drehung des Motors bewegt.Einer der Unterschiede zwischen dieser Installation und ihren früheren Versionen ist das Vorhandensein zusätzlicher Zustände (Schritte): Die frühere Version hatte 4 Zustände und wurde nur durch die Drehschritte des Motors reguliert; Eine neue Option ist ein Sechsgangsystem (Bild Nr. 2).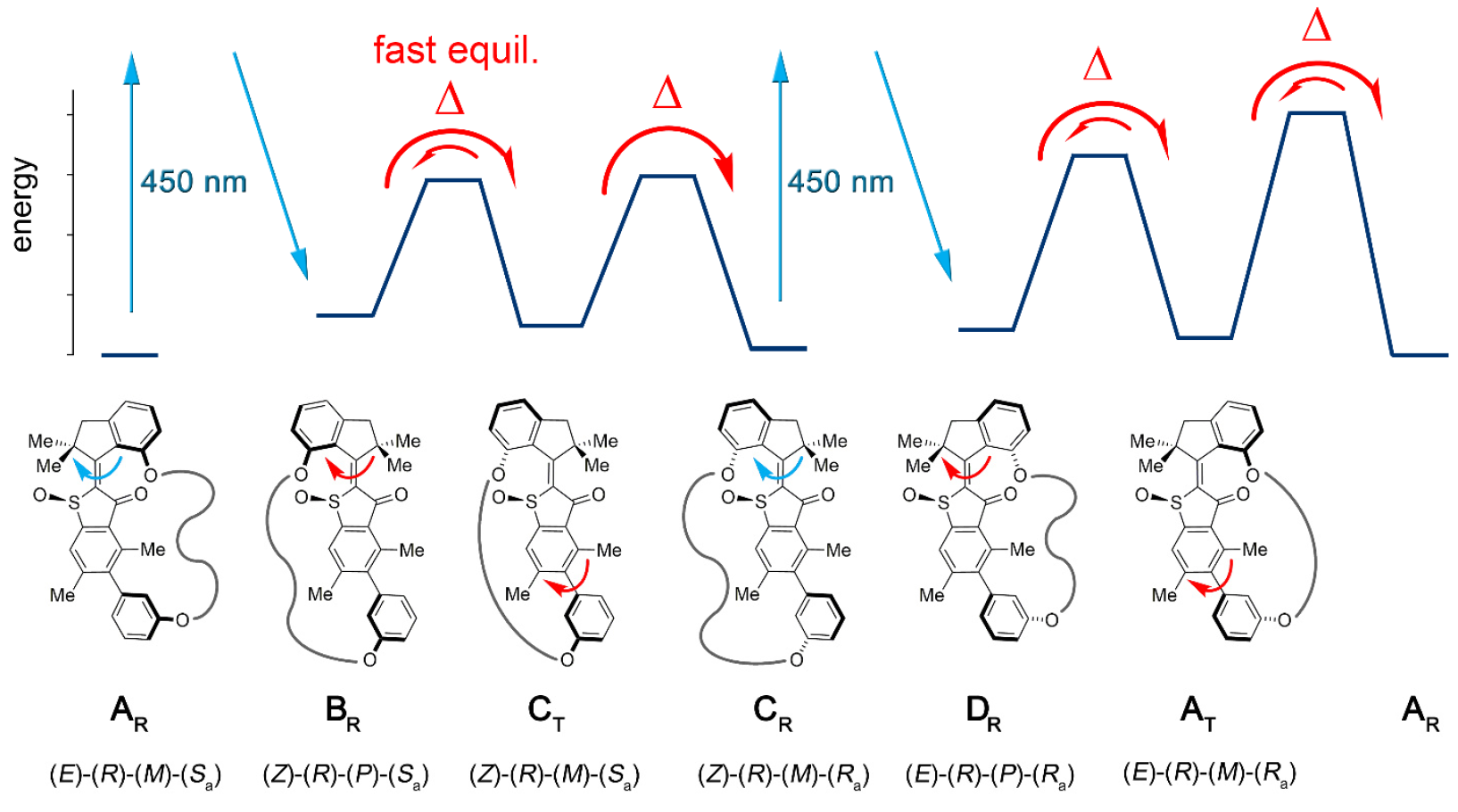 Bild Nr. 2Sechs isomere Zustände werden wie folgt benannt: AT für Spannungszustand A; AR für den entspannten Zustand A sowie BR, CT, CR und DR (Zwischenzustände des Systems).Fünf dieser Schritte können experimentell beobachtet werden, was das Vorhandensein des sechsten Schritts bestätigt und daher die volle Richtwirkung für eine 360-Grad-Drehung des zugehörigen Systems des Blockmotors und des Blockempfängers bestätigt.Das Motorsystem 1 wurde nach dem konvergenten Verfahren aufgrund des bromierten HTI-Vorläufers synthetisiert, an den eine kovalente Bindung mit der Funktion des Boronsäureesters über eine kupferkatalysierte Klickreaktion gebunden war.Anschließend ergibt die Suzuki-Reaktion * eine Makrocyclisierung * , gefolgt von einer Oxidation, um die endgültige Struktur 1 zu ergeben.
Bild Nr. 2Sechs isomere Zustände werden wie folgt benannt: AT für Spannungszustand A; AR für den entspannten Zustand A sowie BR, CT, CR und DR (Zwischenzustände des Systems).Fünf dieser Schritte können experimentell beobachtet werden, was das Vorhandensein des sechsten Schritts bestätigt und daher die volle Richtwirkung für eine 360-Grad-Drehung des zugehörigen Systems des Blockmotors und des Blockempfängers bestätigt.Das Motorsystem 1 wurde nach dem konvergenten Verfahren aufgrund des bromierten HTI-Vorläufers synthetisiert, an den eine kovalente Bindung mit der Funktion des Boronsäureesters über eine kupferkatalysierte Klickreaktion gebunden war.Anschließend ergibt die Suzuki-Reaktion * eine Makrocyclisierung * , gefolgt von einer Oxidation, um die endgültige Struktur 1 zu ergeben.Die Suzuki-Reaktion * ist eine organische Reaktion von Aryl- und Vinylboronsäuren mit Aryl- oder Vinylhalogeniden, die durch Pd (0) -Komplexe katalysiert wird.
Makrocyclisierung * - eine Cyclisierungsreaktion, die zur Bildung eines Makrocyclus führt, d.h. eine chemische Verbindung, in der 9 oder mehr gebundene Atome einen Ring bilden.
Für die beiden stabilsten Zustände von AR und CR von System 1 und den stabilsten Zustand von System 2 wurden für die Strukturanalyse geeignete Kristalle erhalten (®-konfigurierte Isomere von racemischem * AR bei 1d ; CR bei 1e ).Racemat * ist eine äquimolare Mischung zweier Stereoisomere, die Spiegelbilder voneinander sind. Die racemische Verbindung besteht aus Kristallen, in denen sich jeweils Moleküle beider Enantiomere befinden, und ihr Verhältnis beträgt 1: 1.
Da AR / AT und CR mittels HPLC (Hochleistungsflüssigchromatographie) getrennt werden können, kann ihr Verhalten unter Erwärmungs- und Bestrahlungsbedingungen bei unterschiedlichen Temperaturen unabhängig untersucht werden. Wenn die racemische CR-Lösung in (CDCl 2 ) 2 auf 80 ° C bis 140 ° C erhitzt wurde, wurde in 93% der Fälle ein stabileres AR gebildet. Dies legt den Unterschied in der freien Enthalpie ∆G = 1,8–1,9 kcal / mol in diesem Temperaturbereich zwischen diesen beiden Zuständen fest. Eine entsprechende kinetische Analyse ergab eine Hochenergiebarriere von 28,2 kcal / mol bei 80 ° C, die mit der thermischen Isomerisierung der Z / E-Doppelbindung einherging.Beim Abkühlen von reinem CR auf -105 ° C in CD 2 Cl 2 / CS 2(Verhältnis 4/1) und Bestrahlung mit 450 nm Licht erscheint ein neuer Satz von Signalen, der sich von dem bekannten Satz von Signalen AR und AT ( 3a ) unterscheidet. Bild Nr. 3Die Signale des DR-Isomers fallen innerhalb von 28 Minuten bei einer Temperatur von -80 ° C und vollständiger Dunkelheit auf 75% ab, im Gegensatz dazu nehmen die Signale des AT-Isomers zu ( 3 ).Das thermische Gleichgewicht zwischen DR (die verbleibenden 12%) und AT (88%) wird bei -60 ° C beobachtet, was in ∆G = 0,84 kcal / mol zwischen den beiden Zuständen ausgedrückt wird. Die kinetische Analyse der thermischen Zersetzung ergab für diesen Prozess eine begleitende freie Aktivierungsenthalpie ∆ ‡ G = 13,9 kcal / mol bei -80 ° C. Bei Temperaturen von -40 bis 0 ° C werden eine Dämpfung der AT-Signale und ein damit einhergehender Anstieg bekannter AR-Signale beobachtet, bis ein Gleichgewicht zwischen ihnen erreicht ist. Eine kinetische Analyse dieses Prozesses ergab ∆ ‡ G von 18,4 bis 19,3 kcal / mol bei Temperaturen von –40 ... 0 ° C.Somit wurde gefunden, dass die Bestrahlung von CR zum ersten DR-Photoisomerisierungsprodukt führt, das durch Invertieren einer einzelnen Helix im Motorblock thermisch in das AT-Isomer umgewandelt wird. Beim Abkühlen einer Gleichgewichtslösung von AR / AT in CD 2Cl 2 / CS 2 bis -105 ° C und wenn es mit 450 nm Licht bestrahlt wurde, wurde die AR-Photoreaktion aufgrund ihrer Prävalenz beobachtet. Es erscheint auch ein neuer Satz von Signalen, die sich von den bereits bekannten CR ( 3f ) -Signalen unterscheiden . Diese neuen Signale, die das Produkt der AR-Photoreaktion sind, bestätigen die erwartete Richtung der Photoisomerisierung von AR zu BR, die sich dann noch weiter stabilisiert und selbst bei niedriger Temperatur eine schnelle helikale Inversion zu CT erfährt.Bei einer Temperatur von -80 ° C und bei völliger Dunkelheit verschwinden die BR / CT-Signale fast vollständig und nur die CR-Signale verstärken sich weiter ( 3e ). Dies ergibt eine Untergrenze für die Energiedifferenz zwischen CT und CR von 0,98 kcal / mol.Die kinetische Analyse ergab ∆ ‡ G = 13,4–13,5 kcal / mol, was für die Umwandlung von ST in CR erforderlich ist. Es ist ersichtlich, dass die Aktivierungsenergie der Atropisomerisierung von CT in CR im Vergleich zu der für die Atropisomerisierung von AT in AR (18,4 kcal / mol) abnimmt. Dies kann auf die im Vergleich zum AT höhere Spannung (Dehnung) im CT zurückzuführen sein, die auf die großen Abstände zwischen den beiden phenolischen Sauerstoffatomen zurückzuführen sein kann, die als Verbindungspunkte der Linkerkette in den Strukturen C dienen, die sich nicht in den Strukturen A befinden. Im kristallinen Zustand CR zwischen zwei Sauerstoffatomen beträgt 10,6, während es in AR 7,6 ist. Dies gibt der Linkerkette eine größere Konformationsfreiheit (räumliche Position der Atome) in den A-Strukturen als in den C-Strukturen. Berücksichtigen wir die freie Aktivierungsenthalpie für die thermische Umwandlung von CT in CR,dann kann dieser Umwandlungsprozess bei -105ºC vollständig gestoppt werden.Bei einer Temperatur von -105 ° C ist es möglich, den Grad der CT-Akkumulation bei starker Exposition gegenüber AR-Licht zu überprüfen. Dies ist möglich, wenn die Energiedifferenz zwischen BR und ST 2,0 kcal / mol überschreitet und wenn die ST-Struktur selbst nicht an photochemischen Prozessen beteiligt ist. Eine CT-Akkumulation während der AR-Bestrahlung bei -105 ° C ist jedoch nicht möglich, da zwischen CT und BR ein schnelles thermisches Gleichgewicht besteht.
Bild Nr. 3Die Signale des DR-Isomers fallen innerhalb von 28 Minuten bei einer Temperatur von -80 ° C und vollständiger Dunkelheit auf 75% ab, im Gegensatz dazu nehmen die Signale des AT-Isomers zu ( 3 ).Das thermische Gleichgewicht zwischen DR (die verbleibenden 12%) und AT (88%) wird bei -60 ° C beobachtet, was in ∆G = 0,84 kcal / mol zwischen den beiden Zuständen ausgedrückt wird. Die kinetische Analyse der thermischen Zersetzung ergab für diesen Prozess eine begleitende freie Aktivierungsenthalpie ∆ ‡ G = 13,9 kcal / mol bei -80 ° C. Bei Temperaturen von -40 bis 0 ° C werden eine Dämpfung der AT-Signale und ein damit einhergehender Anstieg bekannter AR-Signale beobachtet, bis ein Gleichgewicht zwischen ihnen erreicht ist. Eine kinetische Analyse dieses Prozesses ergab ∆ ‡ G von 18,4 bis 19,3 kcal / mol bei Temperaturen von –40 ... 0 ° C.Somit wurde gefunden, dass die Bestrahlung von CR zum ersten DR-Photoisomerisierungsprodukt führt, das durch Invertieren einer einzelnen Helix im Motorblock thermisch in das AT-Isomer umgewandelt wird. Beim Abkühlen einer Gleichgewichtslösung von AR / AT in CD 2Cl 2 / CS 2 bis -105 ° C und wenn es mit 450 nm Licht bestrahlt wurde, wurde die AR-Photoreaktion aufgrund ihrer Prävalenz beobachtet. Es erscheint auch ein neuer Satz von Signalen, die sich von den bereits bekannten CR ( 3f ) -Signalen unterscheiden . Diese neuen Signale, die das Produkt der AR-Photoreaktion sind, bestätigen die erwartete Richtung der Photoisomerisierung von AR zu BR, die sich dann noch weiter stabilisiert und selbst bei niedriger Temperatur eine schnelle helikale Inversion zu CT erfährt.Bei einer Temperatur von -80 ° C und bei völliger Dunkelheit verschwinden die BR / CT-Signale fast vollständig und nur die CR-Signale verstärken sich weiter ( 3e ). Dies ergibt eine Untergrenze für die Energiedifferenz zwischen CT und CR von 0,98 kcal / mol.Die kinetische Analyse ergab ∆ ‡ G = 13,4–13,5 kcal / mol, was für die Umwandlung von ST in CR erforderlich ist. Es ist ersichtlich, dass die Aktivierungsenergie der Atropisomerisierung von CT in CR im Vergleich zu der für die Atropisomerisierung von AT in AR (18,4 kcal / mol) abnimmt. Dies kann auf die im Vergleich zum AT höhere Spannung (Dehnung) im CT zurückzuführen sein, die auf die großen Abstände zwischen den beiden phenolischen Sauerstoffatomen zurückzuführen sein kann, die als Verbindungspunkte der Linkerkette in den Strukturen C dienen, die sich nicht in den Strukturen A befinden. Im kristallinen Zustand CR zwischen zwei Sauerstoffatomen beträgt 10,6, während es in AR 7,6 ist. Dies gibt der Linkerkette eine größere Konformationsfreiheit (räumliche Position der Atome) in den A-Strukturen als in den C-Strukturen. Berücksichtigen wir die freie Aktivierungsenthalpie für die thermische Umwandlung von CT in CR,dann kann dieser Umwandlungsprozess bei -105ºC vollständig gestoppt werden.Bei einer Temperatur von -105 ° C ist es möglich, den Grad der CT-Akkumulation bei starker Exposition gegenüber AR-Licht zu überprüfen. Dies ist möglich, wenn die Energiedifferenz zwischen BR und ST 2,0 kcal / mol überschreitet und wenn die ST-Struktur selbst nicht an photochemischen Prozessen beteiligt ist. Eine CT-Akkumulation während der AR-Bestrahlung bei -105 ° C ist jedoch nicht möglich, da zwischen CT und BR ein schnelles thermisches Gleichgewicht besteht. Tabelle Nr. 1 Durchdie Kombination der obigen Beobachtungen und Daten können Sie ein mechanisches Bild einer 6-stufigen Sequenz unidirektionaler Rotation in Modell 1 erstellen (Bild Nr. 2 und Nr. 4).
Tabelle Nr. 1 Durchdie Kombination der obigen Beobachtungen und Daten können Sie ein mechanisches Bild einer 6-stufigen Sequenz unidirektionaler Rotation in Modell 1 erstellen (Bild Nr. 2 und Nr. 4). Bild Nr. 4Experimente mit unterschiedlichen Temperaturbedingungen ermöglichten es uns, die Temperaturabhängigkeit von ∆G zwischen CR und AR / AT, DR und AT, AT und AR sowie die Temperaturabhängigkeit von ∆ G für BR / CT und CR, DR und AT, AT und AR zu bestimmen (Tabelle Nr. 1).Im Gegensatz zu dem üblichen 4-Stufen-Mechanismus, der auf dem HTI-Motor basiert, umfasst die vollständige 360-Grad-Drehung zwei zusätzliche Schritte, nämlich die Erhöhung der vom Motor auf den Biarylblock übertragenen potentiellen Energie. Während dieser zwei Schritte wird eine Energiebindung zwischen dem Motor und dem Biarylblock ( 4b ) gebildet, d.h. Letzteres ist nicht mehr energetisch entartet.
Bild Nr. 4Experimente mit unterschiedlichen Temperaturbedingungen ermöglichten es uns, die Temperaturabhängigkeit von ∆G zwischen CR und AR / AT, DR und AT, AT und AR sowie die Temperaturabhängigkeit von ∆ G für BR / CT und CR, DR und AT, AT und AR zu bestimmen (Tabelle Nr. 1).Im Gegensatz zu dem üblichen 4-Stufen-Mechanismus, der auf dem HTI-Motor basiert, umfasst die vollständige 360-Grad-Drehung zwei zusätzliche Schritte, nämlich die Erhöhung der vom Motor auf den Biarylblock übertragenen potentiellen Energie. Während dieser zwei Schritte wird eine Energiebindung zwischen dem Motor und dem Biarylblock ( 4b ) gebildet, d.h. Letzteres ist nicht mehr energetisch entartet. Bild Nr. 5Stattdessen steigt die Energie eines der Atropisomere aufgrund von „Ratschen“ -Schritten des Motors während einer vollen Umdrehung ( 5 s)und 5d ). Diese Energieänderung liefert bis zu 90% der gegenseitigen Umwandlung des Atropisomers in den folgenden thermisch aktivierten Stadien und daher eine fast vollständige Rotation des Biarylblocks während eines Arbeitszyklus des Mechanismus.Diese Beobachtung kann darauf hinweisen, dass der Biarylblock eine Zunahme der potentiellen Energie erfahren hat, die das Ergebnis des Betriebs des Motorblocks ist. Es ist erwähnenswert, dass der anfängliche Anstieg der Energie viel größer ist als der, der den Biarylblock erreicht. Es werden also nur 72% davon vom Motor auf den Biarylblock übertragen.Die obigen Beobachtungen legen nahe, dass die Länge der Verbindung zwischen dem Motor und dem Biarylblock ein wichtiger Aspekt bei der Änderung des Energieübertragungsgrades ist. Daher kann eine kürzere Verbindung ermöglichen, dass mehr potentielle Energie von BR und DR zu AT und CT übertragen wird.Um die Nuancen der Studie genauer kennenzulernen, empfehle ich Ihnen, den Bericht von Wissenschaftlern und zusätzliche Materialien zu lesen.
Bild Nr. 5Stattdessen steigt die Energie eines der Atropisomere aufgrund von „Ratschen“ -Schritten des Motors während einer vollen Umdrehung ( 5 s)und 5d ). Diese Energieänderung liefert bis zu 90% der gegenseitigen Umwandlung des Atropisomers in den folgenden thermisch aktivierten Stadien und daher eine fast vollständige Rotation des Biarylblocks während eines Arbeitszyklus des Mechanismus.Diese Beobachtung kann darauf hinweisen, dass der Biarylblock eine Zunahme der potentiellen Energie erfahren hat, die das Ergebnis des Betriebs des Motorblocks ist. Es ist erwähnenswert, dass der anfängliche Anstieg der Energie viel größer ist als der, der den Biarylblock erreicht. Es werden also nur 72% davon vom Motor auf den Biarylblock übertragen.Die obigen Beobachtungen legen nahe, dass die Länge der Verbindung zwischen dem Motor und dem Biarylblock ein wichtiger Aspekt bei der Änderung des Energieübertragungsgrades ist. Daher kann eine kürzere Verbindung ermöglichen, dass mehr potentielle Energie von BR und DR zu AT und CT übertragen wird.Um die Nuancen der Studie genauer kennenzulernen, empfehle ich Ihnen, den Bericht von Wissenschaftlern und zusätzliche Materialien zu lesen.Epilog
In dieser Arbeit beschrieben Wissenschaftler das Funktionsprinzip ihrer Erfindung, basierend auf der Wechselwirkung von biologischen Motoren und molekularen Elementen. Mit der entwickelten Installation können Sie potenzielle Energie vom Motor auf ein bestimmtes Element übertragen (in diesem Fall handelt es sich um einen Biarylblock). Infolge der motorischen Aktivität muss die thermische Atropisomerisierung von Biaryl unidirektional und schneller ablaufen. Die Energieübertragung erfolgt während thermisch aktivierter Ratschen (Stufen) im Rotationsmechanismus. Es wurde auch gefunden, dass ungefähr 72% der Anfangsenergie des Motors auf eine gegebene Einheit übertragen werden.Diese Erfindung ist ein wichtiger Schritt zur Schaffung vollständiger biologischer Nanometervorrichtungen. Die Verwendung von Biomotoren bei der Entwicklung solcher Mechanismen beruht auf ihrer Biokompatibilität, der einfachen Änderung der Funktionalität aufgrund der Gentechnik sowie ihrer natürlichen Nanoskala. Künstliche Motoren können sich derzeit der oben genannten Kombination von Vorteilen nicht rühmen.Forscher sagen, dass der nächste Schritt in ihrer Arbeit darin bestehen wird, zu demonstrieren, dass ihre Installation nicht nur abstrakte Aktionen ausführen kann, sondern auch nützlich ist. Mit anderen Worten, ihre zukünftige Schaffung wird verfeinert, um nützliche Funktionen auf molekularer Ebene auszuführen.Vielen Dank für Ihre Aufmerksamkeit, bleiben Sie neugierig und haben Sie eine gute Arbeitswoche, Jungs. :) :)Ein bisschen Werbung :)
Vielen Dank für Ihren Aufenthalt bei uns. Gefällt dir unser Artikel? Möchten Sie weitere interessante Materialien sehen? Unterstützen Sie uns, indem Sie eine Bestellung aufgeben oder Ihren Freunden Cloud-basiertes VPS für Entwickler ab 4,99 US-Dollar empfehlen , ein einzigartiges Analogon von Einstiegsservern, das von uns für Sie erfunden wurde: Die ganze Wahrheit über VPS (KVM) E5-2697 v3 (6 Kerne) 10 GB DDR4 480 GB SSD 1 Gbit / s ab 19 $ oder wie teilt man den Server? (Optionen sind mit RAID1 und RAID10, bis zu 24 Kernen und bis zu 40 GB DDR4 verfügbar).Dell R730xd 2-mal günstiger im Equinix Tier IV-Rechenzentrum in Amsterdam? Nur wir haben 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2,6 GHz 14C 64 GB DDR4 4 x 960 GB SSD 1 Gbit / s 100 TV von 199 US-Dollar in den Niederlanden!Dell R420 - 2x E5-2430 2,2 GHz 6C 128 GB DDR3 2x960 GB SSD 1 Gbit / s 100 TB - ab 99 US-Dollar! Lesen Sie mehr über den Aufbau eines Infrastrukturgebäudes. Klasse C mit Dell R730xd E5-2650 v4-Servern für 9.000 Euro pro Cent?Source: https://habr.com/ru/post/undefined/
All Articles